




What You Lose When Your Loading Control Doesn't See What It Claims to See
The undergraduate who loads 20 µg of A549 lysate into a precast gel has been taught a simple rule: run the blot, probe for the protein of interest, strip the membrane, reprobe for β-tubulin, and divide. The ratio tells you whether your target went up or down. The rule is clean enough to fit on a post-it note and wrong enough to have quietly corrupted an unknowable fraction of the published quantitative western blot literature. β-tubulin is not a constant. Its protein abundance shifts with cell cycle phase, tissue type, hypoxia, differentiation state, and drug treatment, and any normalization that treats it as invariant is building a p-value on a moving platform. Worse, the antibody that detects it may be recognizing something else altogether—α-tubulin co-migrating at 50 kDa, a neuron-specific class III β-tubulin isoform upregulated in your cancer model, or a conserved C-terminal epitope shared across the tubulin superfamily that turns your loading control into a tubulin family sum rather than a β-tubulin measurement. You do not know this is happening because the band looks fine. The band always looks fine.
Abbkine's Anti-β-Tubulin Mouse Monoclonal Antibody (3G6), catalog number ABL1030, addresses this problem at the epitope level, and the epitope choice is the specification that separates this antibody from the crowded field of poorly characterized β-tubulin reagents. Clone 3G6 was raised against a synthetic peptide corresponding to the N-terminal twenty amino acids of β-tubulin—MREIVHIQAGQCGNQIGAKF—a sequence that is present in the β-tubulin polypeptide and absent from α-tubulin, γ-tubulin, and all other tubulin superfamily members. The epitope sits in the extreme N-terminal region, upstream of the GTP-binding domain that dominates the conserved core of the protein, in a segment where β-tubulin diverges from its paralogs at the primary sequence level. An antibody that binds this region is not measuring total tubulin. It is not cross-reacting with α-tubulin. It is not blurring the distinction between β-I, β-II, β-III, and β-IV isotypes if those isotypes differ in the N-terminal residues. The signal at 50 kDa on the film is β-tubulin.
The monoclonal architecture of ABL1030 reinforces that specificity. The antibody was produced by hybridoma technology in BALB/c mice, purified from ascites by protein G affinity chromatography, and subjected to validation on 25 or more cell lines that confirmed cross-reactivity with α-tubulin to be below 0.5%. Polyclonal antibodies raised against full-length recombinant β-tubulin can deliver strong signals, but their epitope repertoire shifts from animal to animal and bleed to bleed, and they inevitably contain a fraction of antibodies that recognize conserved domains shared with other tubulins. A monoclonal is a single molecular entity recognizing a single defined epitope, produced by a stable hybridoma clone that will generate the same immunoglobulin molecule for as long as the cell line remains viable. When a reviewer questions whether the loading control normalization in your figure introduced a systematic error, the answer that "the antibody is a monoclonal raised against a β-tubulin-specific N-terminal peptide, with cross-reactivity below 0.5%" is substantially stronger than "the antibody is a polyclonal raised against recombinant protein." The difference between those two sentences is the difference between quantitative western blotting and semi-quantitative western blotting, and reviewers increasingly know the difference.
The species reactivity panel reflects the extraordinary evolutionary conservation of the β-tubulin N-terminus rather than a marketing ambition to list as many organisms as possible. ABL1030 reacts specifically with β-tubulin from chicken, dog, hamster, human, insect, monkey, mouse, rabbit, rat, sheep, and yeast—eleven organisms spanning mammals, birds, insects, and fungi. The inclusion of yeast and insect reactivity is particularly informative: the N-terminal epitope recognized by 3G6 has been conserved across roughly one billion years of eukaryotic evolution, through sequence space that has seen extensive drift in other tubulin domains, because the extreme N-terminus participates in structural contacts that are functionally constrained. For a laboratory that processes western blots from human cell lines, mouse xenograft tissue, and Drosophila or yeast model systems in the same week, this cross-reactivity eliminates the logistical burden of maintaining separate loading control antibodies for each organism. One antibody, one dilution, one band at 50 kDa.
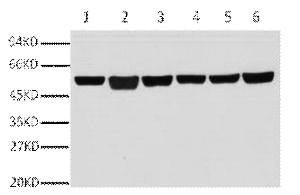
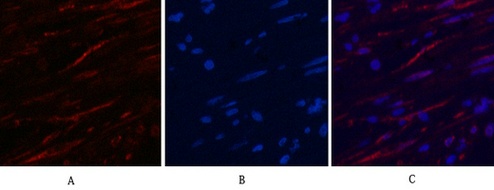
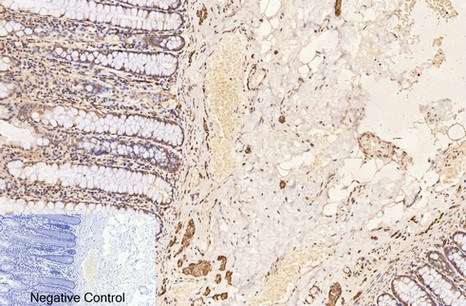
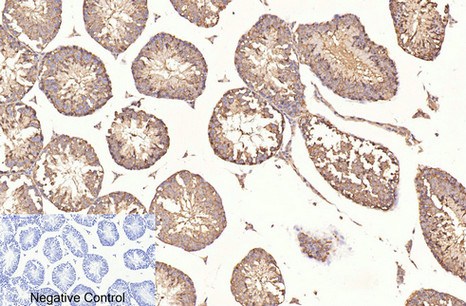
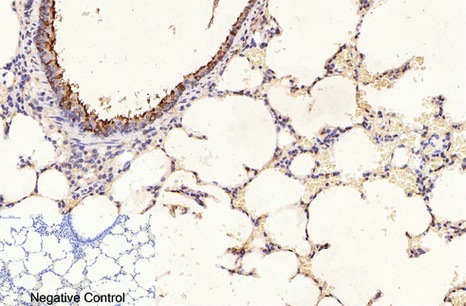
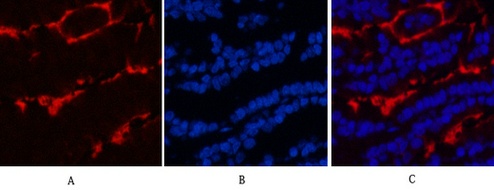
Applications span the three core immunoassay modalities that a loading control antibody must support. Western blot at a recommended starting dilution of 1:10,000, with representative data showing a single clean band in A549 human lung adenocarcinoma cells, rat brain, mouse brain, chicken lung, rabbit testis, and sheep muscle—six distinct tissue types from six distinct species, all producing the same sharp 50-kDa signal without ghost bands, hollow centers, or high-molecular-weight smearing. Immunohistochemistry on paraffin-embedded tissue at 1:400, with demonstration images from human appendix tissue. Immunofluorescence at 1:400, with published representative images in human appendix and mouse lung tissue showing microtubule networks resolved to the level of individual filaments, without the cytoplasmic haze that indicates cross-reactivity with soluble tubulin heterodimers. A loading control antibody that also serves as a cytoskeletal imaging reagent is an antibody that has been validated in two fundamentally different detection contexts—denaturing and native, reduced and cross-linked, linear epitope and conformational epitope—and the fact that 3G6 performs in both contexts tells you something about the accessibility of its N-terminal epitope that a western-blot-only validation cannot.
The publication record is the validation that no manufacturer's certificate of analysis can replicate. ABL1030 has accumulated 23 citations in peer-reviewed literature, a number that places it among the more extensively cited loading control antibodies in Abbkine's catalog. One of those citations appears in the American Journal of Physiology-Lung Cellular and Molecular Physiology, a journal whose reviewers expect quantitative western blot data to be properly normalized and whose editorial standards will not tolerate a loading control that shows batch-to-batch variability or cross-reactivity artifacts. Another citation involves the NPAS3-mediated regulation of VGF expression in neurogenesis and psychiatric disorders—a neuroscience context in which β-tubulin abundance varies across brain regions and developmental stages, and in which an antibody that cross-reacts with neuron-specific β-III tubulin would generate normalization artifacts that could be mistaken for treatment effects. Each of the 23 citations represents an independent laboratory that processed its own samples, ran its own blots, and submitted its own figures to peer review with ABL1030 listed in the methods section. The aggregate signal from 23 laboratories is more informative about real-world performance than any single internal validation dataset.
Formulation and storage specifications reward a practical reading. The antibody is supplied as a liquid solution at 1 mg/mL in PBS, pH 7.4, containing 0.02% sodium azide as preservative and 50% glycerol as cryoprotectant. At 1 mg/mL, a 50-µL vial provides enough material for approximately 5 mL of working solution at the recommended 1:10,000 western blot dilution—enough for roughly five standard minigel membrane incubations at 10 mL per incubation. The 50% glycerol depresses the freezing point, preventing ice crystal formation that denatures immunoglobulin protein during -20°C storage, and the 0.02% sodium azide inhibits microbial growth without the toxicity concerns of higher-concentration azide formulations. Storage instructions specify one-year stability at -20°C from the date of shipment, with centrifugation of the original vial after thawing and prior to cap removal recommended for maximum product recovery, and aliquoting advised to avoid repeated freeze-thaw cycles that progressively reduce antibody binding affinity. These are competent antibody formulation practices stated transparently, and they tell a laboratory manager what they actually need to know: the antibody will survive a year in the freezer, it will not grow contaminants, and it should be aliquoted if the full volume will not be consumed within a few uses.
The question of whether β-tubulin can legitimately serve as a loading control deserves explicit engagement, because the answer is more nuanced than most antibody datasheets acknowledge. The product page itself notes that β-tubulin levels may not be stable in certain cell types, such as adipose tissue—a caution that reflects biochemical reality rather than marketing evasion. β-tubulin expression is upregulated during mitosis when microtubule polymerization increases to accommodate spindle formation. It is downregulated under hypoxia in certain cell lines. It varies across tissue types, with neuronal tissue expressing β-III tubulin at levels far higher than fibroblasts. The appropriate response to this variability is not to abandon β-tubulin as a loading control; it is to verify that β-tubulin abundance does not change under the specific experimental conditions being tested, and to select an antibody whose specificity ensures that the measured signal actually reflects β-tubulin rather than a mixture of tubulin isoforms. A loading control is a measurement, not a constant, and the quality of that measurement is determined by the antibody that generates it. ABL1030 provides a measurement that is monoclonal, epitope-defined, species-validated, application-tested, and publication-vetted.
Several operational notes warrant mention because they reflect the difference between a well-designed antibody and a generic reagent. The recommended starting dilution of 1:10,000 for western blot is a realistic number—the representative images on the product page show clean bands at exactly that dilution across six tissue types, with secondary antibody used at the same 1:10,000 dilution, and no evidence of saturation or signal starvation. The IHC-P and IF recommended dilution of 1:400 reflects the lower antigen density of tissue sections relative to denatured protein bands on a membrane, and the fact that the manufacturer distinguishes between these application-specific dilutions signals that the antibody has been independently optimized for each modality rather than assigned a single dilution that happens to work for western blot. The centrifugation instruction before cap removal prevents loss of antibody that adheres to the cap during shipping, a small detail that can recover 5–10% of the total product for a 50-µL vial. The price point of $59 for 50 µL positions ABL1030 as a cost-effective loading control antibody that does not sacrifice validation depth for affordability.
For the graduate student running their first quantitative western blot, the postdoctoral fellow whose target protein migrates at 42 kDa and cannot use β-actin as a loading control without band overlap, the core facility manager processing blots from three model organisms in a single day, the neuroscientist whose β-tubulin signal must be free of β-III tubulin cross-reactivity, and the cancer biologist whose drug treatment alters microtubule dynamics and needs a loading control whose detection is unaffected by those changes, ABL1030 provides monoclonal specificity, an N-terminal epitope that excludes α-tubulin, eleven-species reactivity, 23 peer-reviewed citations, and a $59 price tag. The loading control that your PI taught you to use in your first week of rotation deserves an upgrade.
Explore full specifications, view representative images, and place your order here: https://www.abbkine.com/product/anti-%ce%b2-tubulin-mouse-monoclonal-antibody-3g6-abl1030/





