




The Polyclonal That Sees What the Monoclonal Misses
There is a quiet truth about the GAPDH loading control market that most manufacturers would prefer to leave unstated. Monoclonal antibodies deliver exquisite specificity—one epitope, one binding site, one signal. That precision is their strength and their limitation. A monoclonal raised against a single peptide sequence will fail to recognize its target the moment that epitope is post-translationally modified, partially degraded, or cross-linked into a protein complex that sterically blocks antibody access. GAPDH is heavily modified in cells: it is S-nitrosylated at Cys152, acetylated at multiple lysine residues, phosphorylated, oxidized, and O-GlcNAcylated, and each of these modifications—many of which are enzymatically removed during standard reducing SDS-PAGE—can ablate the binding of a monoclonal antibody that was raised against an unmodified peptide. When a researcher runs a western blot, transfers the membrane, incubates with a monoclonal anti-GAPDH antibody, and sees a band that should be there but is not, the protein did not disappear. The epitope did. The loading control that was supposed to guarantee equal loading has, instead, guaranteed nothing.
Abbkine's Anti-GAPDH Rabbit Polyclonal Antibody (ABL1021) enters this landscape with a different detection philosophy: instead of targeting a single peptide sequence, it presents a polyclonal repertoire raised against the full-length recombinant GAPDH protein. The result is a pool of antibodies that recognize multiple epitopes distributed across the entire 37-kDa polypeptide—surface-exposed loops, buried hydrophobic patches, and the evolutionarily conserved NAD⁺-binding domain all contributing immunogenic determinants. If one epitope is masked by a residual post-translational modification that survived sample preparation, or by a point mutation in a disease model, or by a splice variant that retains an altered C-terminus, the remaining antibodies in the polyclonal pool still bind and generate signal. That signal redundancy is not a sign of imprecision—it is the feature that keeps the loading control loading under experimental conditions a monoclonal cannot anticipate.
The numbers tell the story of how this redundancy matters in practice. ABL1021 has accumulated 18 publications in peer-reviewed literature, spanning fields from acupuncture neurochemistry to small-molecule drug development for metabolic dysfunction-associated steatohepatitis (MASH). One of those publications, in the Journal of Medicinal Chemistry (impact factor 7.3), relied on ABL1021 for loading control normalization during the preclinical characterization of a silybin derivative targeting hepatic lipid metabolism—an experimental context where GAPDH expression is itself under metabolic regulation and where any antibody that cross-reacts with GAPDH isoforms or pseudogene products would produce normalization artifacts. The study passed peer review in one of the most rigorous medicinal chemistry journals in the world, and the antibody catalog number is visible in the methods section for anyone who cares to verify it.
The biochemical specifications deserve an unhurried examination because they differ from the monoclonal sibling product in ways that are functionally meaningful. ABL1021 was raised in rabbit against recombinant GAPDH protein and then affinity-purified from the resulting antiserum by affinity-chromatography using specific immunogen. The word "affinity-purified" is critical—it separates a reagent that is mostly rabbit IgG with some incidental GAPDH-binding fraction from a reagent that has been enriched for GAPDH-specific antibodies through a column that physically retains only those IgG molecules that bind the immunogen. The resulting product is supplied as a liquid solution at 1 mg/mL in PBS, pH 7.4, containing 0.02% sodium azide preservative and 50% glycerol cryoprotectant, with a recommended starting dilution of 1:10,000 for western blot and 1:200 for immunohistochemistry on paraffin-embedded tissue. The western blot dilution range extends higher or lower depending on sample abundance and detection system sensitivity, as the manufacturer explicitly states that optimal working dilutions should be determined experimentally by the investigator.
Species reactivity is broad and evolutionarily informative. The antibody recognizes GAPDH from chicken, human, monkey, mouse, rabbit, rat, sheep, and Xenopus laevis (African clawed frog). Eight species spanning mammals, birds, and amphibians represents a cross-reactivity profile that is not achieved by accident—it reflects the extraordinary evolutionary conservation of the GAPDH amino acid sequence, which has changed slowly over hundreds of millions of years because the enzyme's glycolytic catalytic function, NAD⁺ cofactor binding pocket, and tetramerization interface are structurally intolerant of variation. For a core facility processing western blots from three different model organisms in a single week, or a research group comparing protein expression between a xenotransplanted human tumor and its murine stromal compartment, this cross-reactivity eliminates the logistical burden of maintaining separate GAPDH primary antibodies for human, mouse, and rat samples.
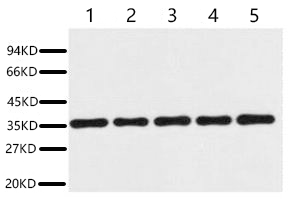
The applications extend beyond western blot in a manner that complements rather than duplicates the monoclonal counterpart. ABL1021 is validated for immunohistochemistry on paraffin-embedded tissue at a suggested starting dilution of 1:200. The product page includes representative western blot images showing a single clean band at 37 kDa in 293T cells, rat brain, NIH 3T3 mouse fibroblasts, sheep muscle, and rabbit testis, diluted at 1:20,000 with secondary antibody also at 1:20,000. The band is sharp, the background is clean, and the signal intensity varies across tissue types in precisely the pattern that GAPDH protein abundance actually follows—highest in metabolically active 293T cells and brain tissue, lower in structural muscle. This is not a loading control that saturates the detector before the target protein becomes visible. It is a loading control that behaves like a protein rather than a constant.
The practical laboratory workflow surrounding ABL1021 benefits from the same thoughtful formulation design evident across Abbkine's antibody product line. The 50% glycerol in the storage buffer depresses the freezing point below -20°C, preventing ice crystal formation that denatures immunoglobulin protein during freezer storage. The 0.02% sodium azide inhibits microbial growth without the toxicity and disposal complications of higher-concentration azide formulations. Storage instructions specify one-year stability at -20°C from the date of shipment, with centrifugation of the original vial after thawing and prior to cap removal recommended for maximum product recovery, and aliquoting advised to avoid repeated freeze-thaw cycles. Shipping occurs on gel packs with blue ice. These are specifications that an experienced immunochemistry laboratory will recognize as competent antibody formulation practice, and that a novice laboratory can follow without specialized knowledge.
The publication record for ABL1021 deserves an analytical, rather than purely promotional, examination. Eighteen publications is a substantial citation count for a loading control antibody, but the meaning of the number changes when one inspects the journals in which those citations appear. The Ziwuliuzhu acupuncture study deposited on Research Square used the antibody in a rat model of insomnia, demonstrating the functional conservation of the GAPDH across experimental animals. The Journal of Medicinal Chemistry citation placed the antibody in a high-impact medicinal chemistry context, where reviewers scrutinize the normalization of quantitative western blots with a rigor that general biology journals rarely match. Each of the remaining sixteen publications represents an independent laboratory that prepared whole-cell lysates, separated proteins by SDS-PAGE, transferred to membrane, incubated with ABL1021, exposed the blot, and looked at the resulting film or digital image—and concluded that the band at 37 kDa was clean enough, specific enough, and single enough to serve as a loading control in a figure that a reviewer would eventually examine.
The broader biological context of GAPDH as a moonlighting protein adds a dimension to antibody selection that most loading control datasheets ignore. GAPDH catalyzes the sixth step of glycolysis—the oxidative phosphorylation of glyceraldehyde-3-phosphate to 1,3-bisphosphoglycerate—and it has performed this function in eukaryotic cells for roughly two billion years. But GAPDH is also one of the most promiscuous moonlighting proteins ever characterized: it translocates to the nucleus upon S-nitrosylation and participates in transcriptional regulation of the apoptosis program, it binds to the 5' untranslated regions of specific mRNAs and modulates their translation, it associates with the cytoplasmic face of the endoplasmic reticulum and is required for ER-to-Golgi vesicular trafficking, it contains a peptide sequence with intrinsic antimicrobial activity against E. coli, P. aeruginosa, and C. albicans, and it has been reported to act as a transferrin receptor on the macrophage cell surface. The same 37-kDa band that researchers normalize against in a western blot is simultaneously a glycolytic enzyme, an apoptosis regulator, an mRNA-binding protein, and an antimicrobial peptide precursor. The antibody that detects GAPDH as a loading control is also, in a different experimental design, the antibody that could detect GAPDH in its moonlighting roles. A polyclonal recognizing multiple epitopes distributed across the entire polypeptide is better positioned to capture GAPDH in its various post-translationally modified and conformationally distinct states than a monoclonal recognizing a single linear epitope that may be unavailable when the protein is bound to nucleic acid or incorporated into a multiprotein complex.
The monoclonal sibling product ABL1020—discussed in its own dedicated profile—offers batch-to-batch consistency and defined epitope specificity. ABL1021 offers epitope redundancy and the signal stability that arises from polyclonal recognition of multiple determinants on the same target. The choice between monoclonal and polyclonal for a loading control antibody is not a choice between good and bad. It is a choice between two forms of reliability—one achieved through precise epitope definition and hybridoma stability, the other achieved through recognition breadth that insures against epitope loss. Laboratories processing samples whose GAPDH may carry genetic variants, whose tissue preparation protocols may partially degrade the protein, or whose experimental treatments may induce transient post-translational modifications that persist through sample processing will find that ABL1021's polyclonal architecture provides a margin of safety that a monoclonal cannot offer.
Several usage notes merit explicit mention because they reflect operational immunochemistry discipline rather than marketing language. The recommended starting dilution of 1:10,000 for western blot is a realistic number—not a marketing claim that requires four-hour exposures to see any signal, and not an over-concentration that saturates the band on a standard 30-µg lysate load. The validation data on the product page show clean bands at 1:20,000, suggesting that the 1:10,000 recommendation leaves headroom for weaker expression samples. The IHC-P starting dilution of 1:200 is informed by the fact that paraffin-embedded tissue sections present a lower antigen density than denatured protein bands on a membrane, and antibodies validated at 1:10,000 for western blot will not necessarily work at 1:10,000 for IHC—the protocol reflects this distinction. The centrifugation instruction before cap removal prevents loss of product that adheres to the cap during shipping and handling. The aliquoting recommendation prevents freeze-thaw cycling that progressively denatures immunoglobulin protein, a phenomenon that is real, measurable by ELISA, and responsible for the slow decline in western blot signal intensity that many laboratories attribute to membrane transfer efficiency because they have never tracked antibody age as an experimental variable.
For the laboratory that runs western blots on tissue from multiple species and cannot maintain separate GAPDH primary antibodies for each organism, for the investigator whose experimental treatment conditions may induce partial GAPDH degradation or modification that ablates a monoclonal epitope, for the core facility that processes blots from diverse biological sources and needs a loading control antibody that generates a clean single band at 37 kDa regardless of sample origin, and for the researcher who understands that a loading control is not a constant but a measurement of a protein whose abundance can shift with treatment and whose epitope availability can change with modification state, ABL1021 provides polyclonal recognition breadth, eight-species reactivity, 18 peer-reviewed publication citations, affinity-purified specificity, and a formulation stable at -20°C for one year. The polyclonal that sees what the monoclonal misses is available in 50 µL and 200 µL vial sizes, ships on blue ice, and stores indefinitely at -20°C with proper aliquoting.
Explore full specifications, view representative data, and place your order here: https://www.abbkine.com/product/anti-gapdh-rabbit-polyclonal-antibody-abl1021/





