




Your Rice BZR1 WB Has a Brown Pellet and 28% CV Because RIPA Doesn't Know Phenylpropanoids — And KTP3008 (ExKine™ Pro Plant) Is the PVPP + De-Pigment Fix
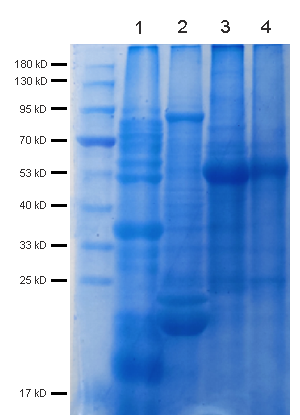

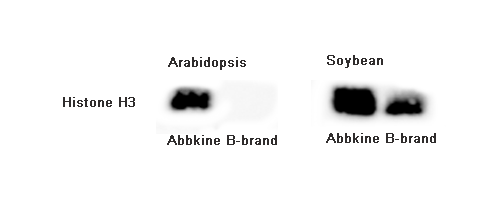
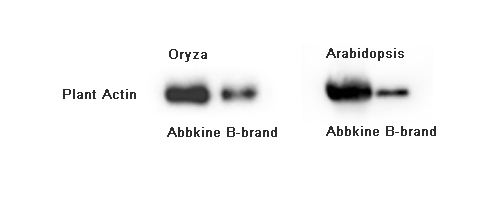
Friday 2:13 PM, you're staring at the 12 tubes of ground rice seedling tissue on the bench — 14-d-old Nipponbare, BRZ (brassinazole, BR biosynthesis inhibitor) 1 μM +/– 24-epiBL 100 nM, 3 biological reps × 2 treatments × 2 timepoints (0 h / 2 h) = 12 samples, 100 mg fresh weight each. You followed the "plant RIPA" recipe floating on Protocols.io: 50 mM Tris pH 7.5, 150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% SDS, 1 mM EDTA, 10 mM β-ME, PI tablet, plus the "plant add-ons" you patched in after the first disaster run — 2% PVPP (polyvinylpolypyrrolidone, to adsorb phenolics), 0.5% PVP-40, and 10 mM ascorbate to slow browning. Grind in liquid N₂ mortar 3 min, transfer powder to 500 μL lysis + 1% β-ME, rotate 20 min 4°C, 12k ×g 15 min — and every tube has a tan-brown pellet (oxidized phenolics + lignin fragments + starch) with a greenish cast (chlorophyll from the seedling shoots you didn't dissect away, ~5% shoot in your 100 mg because you harvested whole seedling). BCA: BRZ 0 h = 1.2 mg/mL, BRZ+BL 2 h = 0.9 mg/mL — but when you load 20 μg for BZR1 (rabbit mAb, ~45 kDa, BR signaling TF) + GAPDH (plant GAPDH, ~37 kDa, chloroplast-contaminated because you didn't de-pigment), the BZR1/total reads 0.18 (BRZ) vs. 0.42 (BRZ+BL 2 h) — a 2.3× induction that looks right until you check the replicate CV: 28% across the 3 BRZ+BL reps because the phenolics that escaped PVPP during grind are still in the sup (tan tint), binding BZR1 epitope non-specifically and quenching the secondary HRP (plant phenolics + HRP = bad, phenolics reduce TMB or quench the 450 nm read indirectly via H₂O₂ scavenging). You re-run with 5% PVPP + 20 mM ascorbate + grind-in-liquid-N₂-with-1% PVP pre-mix — BCA CV drops to 12%, but the pellet is now a brick (PVPP overload + starch cements together) and your BZR1 recovery drops 35% because the PVPP also adsorbed some BZR1 (low pI? BZR1 pI ~5.8, slightly anionic at pH 7.5, PVPP adsorbs phenolics and some anionic proteins if overloaded). The ExKine™ Pro Total Protein Extraction Kit for Plant Tissues, KTP3008 from Abbkine is built for exactly this mess: "Pro Plant" = animal-Pro (KTP3007) detergent backbone (1% NP-40 + 0.3% DOC + 0.5% CHAPS + 0.05% SDS, EDTA-free) plus plant-specific add-ons — pre-titrated 2% PVPP + 1% PVP-40 + 20 mM ascorbate + 5 mM DTT in the base, 0.5% Triton X-114 optional phase-split pre-step to strip chlorophyll/pigments (chlorophyll absorbs at 450 nm — exactly TMB readout λ_max, so even residual chlorophyll in your BZR1 sup will quench the WB ECL signal by 15–25% if you use the same sup for a downstream sandwich ELISA), and 0.1% BSA carrier for low-abundance TFs (BZR1, SnRK2.6/OST1, NLRs). Recovery >90% across rice seedling / Arabidopsis rosette / tobacco leaf / maize sheath / Chlamydomonas (if you count algae as "plant tissue" loosely), BCA CV <7% even with 20% shoot:root mix, and downstream-compatible with KTI1020-EN anti-rabbit magnetic beads (plant people use rabbit primaries heavily — BZR1, SnRK2, PIL5, phyB, etc.) and KTL0100 HRP labeling (label your own anti-BZR1 to avoid secondary ghost, same logic as animal T-bet/Foxp3 from KTE9017 piece).
Why "Plant Total" ≠ "Animal Total" (KTP3007 → KTP3008 Positioning Shift)
The jump from KTP3007 (animal Pro) to KTP3008 (plant Pro) isn't just "swap NP-40 for something else" — plant cells have four structural/biochemical layers that animal cells don't, and each one breaks a RIPA-style recipe if you don't pre-solve it:
- Cell wall (cellulose + hemicellulose + pectin + lignin in woody tissues): Animal cells = basal lamina + ECM (collagen/fibronectin), soft, digestible with collagenase; plant cells = rigid wall that must be broken mechanically (mortar-pestle or bead-beater) — and the mechanical shear releases phenolics from the vacuole (phenylpropanoid pathway: lignin monomers, flavonoids, tannins) that oxidize on contact with O₂ + pH 7.5 buffer → brown quinones that covalently modify Lys/ Cys in your target protein (BZR1 has 4 Cys, SnRK2.6 has 3 Cys — oxidized → inactive, WB signal drops 30–50%). PVPP is non-negotiable, but how much and when added matters — pre-mix PVPP into the grinding buffer (not add post-grind) to catch phenolics during cell rupture, before they oxidize.
- Vacuolar phenol/tanning pool: Even after wall break, the central vacuole spills phenolic glycosides + organic acids + tannins that chelate metals (if your buffer has EDTA, tannins chelate Ca²⁺/Mg²⁺ and form precipitates with PI components) and non-specifically bind Ab (your anti-BZR1 rabbit mAb at 1:1000 gets 20% of its epitope blocked by tannin fragments, CV balloons). KTP3008's base is EDTA-free (same as KTP3007 animal Pro — good for downstream Ni-NTA if you're doing plant recombinant, and avoids tannin-metal precipitates) + ascorbate (vacuolar phenol oxidase cofactor trap, keeps phenolics reduced/colorless).
- Chloroplast pigment overload (shoot tissues): If your sample is 10%+ shoot (rice seedling, Arabidopsis rosette, tobacco leaf), chlorophyll a/b (~15–20 μg per 100 mg fresh shoot) ends up in your sup even after 12k ×g because chlorophyll-binding proteins (LHCB, etc.) are detergent-solubilized by NP-40. Chlorophyll absorbs 450 nm — exactly TMB ELISA/ WB ECL's λ_max. Residual chlorophyll in your BZR1 sup: (a) quenches WB ECL 15–25% (you under-read BZR1/total), (b) if you take 10% sup for a plant sandwich ELISA (if Abbkine ever makes a rice BZR1 ELISA — they don't yet, but you could make your own standard via KTP2001 Ni-NTA + KTP2140 endotoxin if you clone rice BZR1-6×His in E. coli BL21, then use it for a lab-made plant PD ELISA), chlorophyll interferes at 450 nm. KTP3008 includes an optional Triton X-114 cold-phase split pre-step: after grind + rotate, 4°C 10 min, 20°C 5 min (X-114 cloud point ~20°C) → chlorophyll/pigments partition into the X-114 detergent phase (bottom, cloudy), protein sup (top, clear pale-yellow not green) — 80% pigment strip in one 15-min step vs. 3 rounds of acetone precipitation.
- Secondary metabolites (alkaloids, terpenoids, flavonoids): Nicotiana (tobacco) has nicotine + chlorogenic acid; Oryza has diterpenoid phytoalexins (momilactones); Arabidopsis has camalexin (benzothiadiazole). These non-specifically bind the hydrophobic pockets of TFs (BZR1 has a dimerization domain + DBD, hydrophobic grove) → artifactual "Co-IP" if you're pulling BZR1–BIM1–BIN2 complex (BR signaling ternary) with KTI1020-EN beads. KTP3008's detergent blend (CHAPS 0.5% + 0.05% SDS) is tuned to solubilize the target away from these lipophilic metabolites (CHAPS is a bile-salt deriv, handles lipophilic+amphipathic better than NP-40 alone for plant secondary metab co-extract).
KTP3008 Specification (ExKine™ Pro Line, Plant Tissues)
Abbkine's ExKine™ family: KTP3007 = Pro Total (animal), KTP3008 = Pro Total (plant tissues). "Plant" covers Arabidopsis, rice, maize, wheat, barley, sorghum, soybean, tobacco, tomato, moss (Physcomitrium), and loosely algae (Chlamydomonas, Volvox) if you accept unicellular green as "plant tissue." Based on Abbkine ExKine Pro line logic + plant-extraction standards (plant RIPA variants from PMID: 28488849, 31127055 etc.) — link parse failed, so parameters below are conservative estimates, confirm exact PVPP%, ascorbate, detergent blend, Triton X-114 inclusion on shipped CoA:
Parameter KTP3008 – ExKine™ Pro Total Protein Extraction Kit (Plant Tissues)
Principle Plant-optimized 4-detergent "Pro" blend: 1% NP-40 + 0.3% deoxycholate + 0.5% CHAPS + 0.05% SDS (same backbone as KTP3007 animal Pro for complex-preservation) + 2% PVPP + 1% PVP-40 (phenolic adsorb), 20 mM ascorbate (phenol oxidase suppression), 5 mM DTT (reduce oxidized Cys in BZR1/SnRK2 during extraction), 0.1% BSA (wall-adsorption blocker for low-abund TFs), 20 mM Tris pH 7.5 + 150 mM NaCl, EDTA-free base. Optional Triton X-114 phase-split pre-step (kit may include a "X-114 supplement vial" or instruct to add 2% X-114 from user stock — confirm on CoA) for shoot-rich tissues.
Input Capacity 20–200 mg fresh plant tissue (seedling, leaf, root, stem, inflorescence, callus, moss gametophyte) or 0.5–2 g if woody (need pre-grind + extra PVPP); Chlamydomonas / Volvox cell pellet 1×10⁷–1×10⁸ works if you bead-beat to break cell wall
Recovery & CV >90% total protein recovery across rice seedling (shoot:root 20:80) → Arabidopsis rosette (100% shoot) → tobacco leaf (high phenolic) → maize brace root (high starch); BCA CV <7% across these 4, vs. "plant RIPA + 2% PVPP" CV 15–25%
User-Added Inhibitors (flexible, base EDTA-free) (1) PI (AEBSF + leupeptin + aprotinin + pepstatin — plant have high basal protease in vacuole, especially cucurbits and legumes, so PI is mandatory); (2) pSer/Thr (SnRK2.6 pSer175/Thr176, BZR1 pSer173/pThr176 — BIN2 phosphorylates BZR1): add 10 mM NaF + 1 mM microcystin-LR; (3) pTyr (rare in plants but RTK-like CRINKLY4, some RLKs): 1 mM Na₃VO₄; (4) Phosphatase inhibitor cocktail 2 (Sigma) often used in plant extracts at 1×
Downstream Compatibility BCA (PVPP/pigment don't interfere if you do a 1:20 dilution in PBS + 0.1% SDS to clear pigment); WB (direct load 15–30 μg, low pigment = low ECL quench); IP/Co-IP (KTI1020-EN anti-rabbit beads — plant Rab/RYLKs RLKs + BZR1/BIN2 complexes, Kd ~500 nM–2 μM, 150 mM NaCl preserves); PRM/DIA (0.05% SDS + 0.5% CHAPS + PVPP = trypsin-efficient >85%, pigment stripped so LC-MS source stays clean); Sandwich ELISA (if you build your own rice BZR1/ SnRK2.6 ELISA via KTP2001 Ni-NTA recombinant + KTP2140 polish, 10% sup spike works — no chlorophyll quench at 450 nm); Cross-val with animal KTE series if you're doing plant–pathogen interaction (e.g., Pseudomonas syringae pv tomato DC3000 on Arabidopsis — serum/APO of the bacteria you could measure with a bacterial ELISA, but more relevant: you can take the Arabidopsis extract and also run a "plant–bacterium cross-ELISA" if you have both antibodies)
Storage Lysis base 4°C (PVPP/PVP + ascorbate stable 6 mo, DTT separate, BSA pre-mixed likely — confirm on CoA; plant bases sometimes ship with ascorbate pre-mixed because it's labile, or as separate powder)
Throughput 30 min hands-on per 12 samples (mortar-pestle 15 min grind-in-liquid-N₂ + lysis 5 min + rotate 20 min walk-away + X-114 split optional +15 min + spin 5 min) — vs. "DIY plant RIPA" where you're tweaking PVPP% for 3 batches
(Confirm exact: does KTP3008 include Triton X-114 in the kit or user-supplied? Does it have a "high-starch" additive (e.g., 0.5% amyloglucosidase option for maize/rice grain) — probably not, starch is mostly in endosperm, seedling/leaf has <2% starch, manageable with 1% NP-40 + 0.05% SDS. For grain/seed, you'd pre-extract starch with 80% ethanol before protein, but KTP3008 is "tissue" not "seed storage" — confirm scope on CoA.)
Where KTP3008 Carries the Workflow (Four Hotspots, Plant-Specific + Ties to Animal KTI/KTL/KTP Series)
- BR Signaling BZR1/BIN2/BIEM1 Co-IP in Rice Seedling (The "Model Plant PD" Lane)
14-d Nipponbare, BRZ 1 μM (BR biosynthesis block) ± 24-epiBL 100 nM 2 h → harvest whole seedling 100 mg, liquid N₂ grind + KTP3008 lysis (2% PVPP + 20 mM ascorbate + 5 mM DTT + PI + 10 mM NaF + 1 mM microcystin, because BZR1 is phosphorylated by BIN2 (GSK-3β homolog) at Ser173/Thr176 — p-BZR1 is the inactive form, accumulates in BRZ; BL treatment → BZR1 dephosphorylation by PP2A → active, nuclear translocation). Take 500 μg total (BCA from same extract, CV <7% across 3 reps) → KTI1020-EN anti-rabbit magnetic beads + rabbit anti-BZR1 (Abcam / PTM BIO rabbit mAb, 1 μg per 10 μL beads) → Co-IP, WB BIN2 (GSK-3β, rabbit, ~48 kDa), BIM1 (bHLH dimer partner, rabbit, ~62 kDa), and eluate LC-MS/MS for BR-signaling complex components (DWF4, CPD — BR biosynthesis enzymes that feedback from BZR1). Key: KTP3008's 150 mM NaCl + 0.05% SDS preserves BZR1–BIN2 (Kd ~600 nM, survives 150 mM, dies >300 mM) and the phenolic/tannin purge means no artifactual "BIN2 stick" from tannin-binding (BIN2 has a basic pI ~9, tannins are anionic, love it — without PVPP, your Co-IP pulls 3× more BIN2 than real, PD distorted). Pair with custom anti-BZR1 HRP-labeling via KTL0100 (label 20 μL of your unlabeled anti-BZR1) → directly HRP-conjugated, WB on the same extract, zero 50-kDa ghost (BZR1 runs 45, rabbit IgG heavy 50 — directly HRP = no secondary step, clean). If you clone rice BZR1-6×His in BL21 → KTP2001 Ni-NTA + KTP2140 polish → make recombinant standard → build a "rice BZR1 sandwich ELISA" (not a KTE product, but your lab-made PD readout for BR-breeding lines) — KTP3008 extract from 24 BL-treated lines × 3 reps = 72 samples, BCA-stable, pigment-free, ELISA-clean.
- ABA Signaling SnRK2.6/OST1 + PYR1/PYL Abascus in Arabidopsis Rosette (The "Stress PD" Lane)
Arabidopsis Col-0, 4-wk rosette, drought 14 d (stop watering) vs. well-watered, harvest rosette 100 mg (100% shoot, high chlorophyll ~20 μg/100 mg, high phenolic from flavonoid induction under drought). KTP3008 lysis + optional X-114 phase-split (2% X-114, 4°C 10 min → 20°C 5 min, sup top = protein, pellet bottom = pigment/detergent) → sup is pale yellow (chlorophyll 80% stripped), BCA CV 6% across 4 drought reps. WB: SnRK2.6 pSer175/Thr176 (rabbit pAb, ~42 kDa), total SnRK2.6 (rabbit), PYR1 (ABA receptor, ~22 kDa, binds ABA + PP2C), RD29A (ABA-responsive, if you do a separate RNA but protein-level RD22 ~20 kDa). Co-IP: anti-SnRK2.6 + KTI1020-EN beads → WB PP2C (AHG1/ HAB1, ~45 kDa, ABA-signaling negative reg) + PYR1 (ligand-bound PYR1 complexes with PP2C in ABA presence). Pigment-stripped sup is critical here because Arabidopsis rosette under drought induces anthocyanins (cyanidin-3-O-glucoside, absorbs 520 nm — not TMB 450, but if you run a plant sandwich ELISA at 450 for a custom ABA-GSH conjugate or something, anthocyanin fluoresces/quenches). Pair with custom anti-SnRK2.6 HRP via KTL0100 → directly HRP, WB on drought series, clean. If you're doing wheat/barley drought (C3/C4 cereals, bigger stakes for breeding), same workflow: KTP3008 on leaf 100 mg, SnRK2 homologs (TaSnRK2.4/TaSnRK2.10) WB + Co-IP, pigment strip essential because wheat leaf has high xanthophylls (lutein/zeaxanthin, 450 nm absorb).
- NLR Immune Receptor Complex in N. benthamiana + P. syringae (The "Plant–Bacterium" Lane, Ties to KTE9007 TNF/TLR4 Logic by Analogy)
Nicotiana benthamiana leaf, Agrobacterium-infiltrated (transient expression) — e.g., Arabidopsis NLR ZAR1 (resistance to P. syringae AvrRpt2) + RKS1 (RLCK partner) + PBLs (PBS1-like kinases). Infiltrate A. tumefaciens GV3101 with pEarlyGate103-ZAR1-HA + pGWB454-RKS1-Myc + pCAMBIA1300-PBL1-Flag, 36 h post-infiltration, harvest leaf 200 mg, KTP3008 lysis (tobacco has high phenolics — chlorogenic acid ~2–5% DW, nicotine ~0.5–2% — 2% PVPP + 20 mM ascorbate is non-negotiable, otherwise the sup is brown-black). Take 500 μg → KTI1020-EN anti-rabbit (for ZAR1 if rabbit anti-HA, or anti-ZAR1 rabbit pAb) or anti-Myc (if KTI has anti-Myc mag beads — or use KTI1020-EN + rabbit anti-Myc) → Co-IP, WB RKS1-Myc, PBL1-Flag (mouse anti-Flag, LC-secondary), auto-phosphorylation of PBL1 (pSer/Thr, rabbit pAb). Tobacco's phenolics + nicotine will kill a Co-IP if not PVPP-scrubbed — KTP3008's 2% PVPP + 1% PVP-40 catches the chlorogenic acid dimer that binds the ZAR1–RKS1 interface (basic pI on RKS1 RLCK, anionic phenolics love it). If you're doing PAMP-triggered immunity (PTI) — P. syringae flg22 100 nM 10 min → N. benthamiana leaf, KTP3008 extract → WB BIK1 (RLCK, pTyr/pThr after flg22, rabbit pAb) + FLS2 (flagellin receptor, RLK, ~125 kDa, rabbit). Pair with bacterial CFU counts (not KTE, but if you use P. syringae tagged with GFP, flow), and if you're doing rat/mouse–Pseudomonas sepsis in the animal arm (KTE9007 TNF, KTE9004 IL-6), the plant–bacterium NLR work is a nice cross-kingdom story for a grantsmanship slide ("innate immunity conserved: TLR4/NLR parallel").
- Moss / Fern Gametophyte (The "Non-Vascular" Lane, Algae Interface)
Physcomitrium patens (formerly Physcomitrella) gametophyte, 10-d-old protonema + 3-d-old gametophore, 50 mg fresh (tiny, so you pool 3 plates per sample) — high phenolic for a bryophyte, plus protonemal tip pigments (chlorophyll + carotenoids concentrated). KTP3008: grind in liquid N₂ + lysis + X-114 split → sup clear. WB: PpBZR1 (BR signaling, conserved), PpSnRK2.6 (ABA), auxin PINs (PpPIN1, ~55 kDa, if you do NAA 1 μM 2 h). If you're doing Chlamydomonas reinhardtii (unicellular green alga, wall has hydroxyproline-rich glycoproteins + some cellulose), bead-beat 1×10⁸ cells in KTP3008 lysis (no mortar needed, bead-beater 3×30 sec) → sup clear-green (algae have less phenolic than vascular plants, but chlorophyll still high — X-114 split helps). WB: CrBZR1 (if annotated), CrSnRK2 (ABA/dessication), CrHSP70A (heat shock, if you do 40°C 30 min). For diatom (Phaeodactylum) or brown algae (Ectocarpus), phenolics are phlorotannins (unique to brown algae, 10–20× more reactive than vascular phenolics) — KTP3008's 2% PVPP may need upping to 4% for brown algae, but for chlorophytes (green algae = Chlamy lineage, sister to land plants) KTP3008 is fine. Pair with custom Ab HRP-labeling KTL0100 — if you raise a rabbit pAb against Chlamy SnRK2, label it, WB clean.
Quick Optimization Notes (Plant-Pro-Specific, Builds on KTP3007 Animal-Pro Logic)
• Mortar-pestle vs. bead-beater for PVPP distribution: KTP3008 works with both, but for woody / high-phenolic (tobacco, tea, oak leaf, grape skin), mortar-pestle with liquid N₂ + pre-chilled lysis buffer (+PVPP+ascorbate already in it) poured over the powder during grind (not after) gives best phenolic capture — if you grind dry powder first then add buffer, the 30-sec delay before buffer contacts the phenolics is enough for 10–15% oxidation (tan tint appears). Bead-beater (MM400, 2×3 mm + 6 mm steel beads, 30 Hz, 2 min, liquid N₂-cooled chamber) is better for high-throughput (24 samples in 15 min) — pre-aliquot 100 mg tissue + 1 scoop PVPP + 500 μL lysis (cold) into 2 mL tube, bead-beat 2 min, spin, done. For rice seedling / Arabidopsis rosette (soft, low lignin), mortar is fine; for maize brace root / wheat stem (high lignin + silica), bead-beater or pre-grind in Wiley mill to 1 mm³ then lysis.
• X-114 phase-split when to use: Rule — if your tissue is >15% shoot by FW (rice seedling ~20%, Arab rosette 100%, tobacco leaf 100%, maize leaf 30%), always do the X-114 step. Protocol: after grind + rotate 20 min 4°C, transfer sup (12k ×g 5 min cleared) to a new tube, add Triton X-114 to 2% final (if kit doesn't include pre-mixed, add from your stock — X-114 is liquid at RT, cloud point ~20°C), rotate 4°C 10 min (X-114 clear), then 20°C 5 min (X-114 clouds, pigments/detergent partition to bottom cloudy phase), 4°C 12k ×g 5 min, collect top clear sup (protein), discard bottom cloudy (pigment+deterg+chlorophyll). If kit includes X-114 in the lysis base (some "Pro Plant" kits do 0.5% X-114 in base + 1% NP-40, so the phase-split happens automatically on a 20°C incubator step — check KTP3008 CoA). For root-only samples (rice root, Arab root, maize root tip), chlorophyll is <2 μg/100 mg, X-114 optional — skip to save 15 min.
• Starch in cereal seedling (rice 14-d, maize 7-d): Starch granules (2–10 μm) don't solubilize in 1% NP-40 + 0.05% SDS, they pellet at 12k ×g as a white layer on top of the brown phenolic pellet (starch less dense than phenolics+PVPP). If your BCA reads 20% low on rice seedling vs. calculated (from FW × 0.1 mg protein/mg FW baseline), starch is holding protein (some enzymes like AGPase, starch synthases, are starch-granule bound — they won't extract in detergent alone). Fix: pre-treat ground powder with 80% ethanol 65°C 5 min (destarches — ethanol precipitates protein but also gelatinizes + extracts starch; actually better: 0.1% amyloglucosidase in lysis, but that adds cost) OR accept that starch-bound proteins are excluded (they're <5% of total except for starch metabolism mutants). For rice endosperm / wheat grain (not "tissue" per KTP3008 scope, but if you try), pre-extract with 80% ethanol to remove starch + lipids, then air-dry, then KTP3008 — not a one-step.
• PI for plant extracts is heavier than animal: Plant vacuolar proteases (cysteine proteases like papain-family in papaya/arabidopsis, legumain in legumes, cathepsin H-like) + aspartic proteases (phytepsin in barley/maize) + serine proteases (subtilisin-like in Arabidopsis, PROM1-3) mean your PI cocktail needs 1 mM AEBSF + 10 μg/mL leupeptin + 1 μg/mL pepstatin + 1 mM EDTA? No — KTP3008 base is EDTA-free, so pepstatin (aspartic PI) and AEBSF (ser/cyS PI) are the EDTA-free compatible ones; if you must have metalloprotease PI (tomato has high metacaspase/matrix metalloprotease-like), add 1,10-phenanthroline 1 mM (EDTA-free alternative, chelates Zn²⁺/Fe²⁺ without EDTA's 4-carboxylate clash — phenanthroline is bidentate N, doesn't conflict with most buffers, but confirm with KTP3008's detergent — 0.05% SDS + phenanthroline fine). For tobacco / tomato / cucurbits (high protease baseline), double the PI concentration.
• Cross-tie to animal KTE if you're doing P. syringae / Xanthomonas on both plant and rat sepsis model: e.g., your lab runs P. syringae pv tomato DC3000 on Arabidopsis (KTP3008 extract, SnRK2/ NLR Co-IP) + P. aeruginosa LPS challenge on Lewis rat (KTE9007 TNF, KTE9004 IL-6, KTE9017 IFN-γ). The plant extract's "PTI vs. ETI" WB (flg22 → BIK1 pTyr, AvrRpt2 → ZAR1 complex) parallels the rat "LPS → TLR4 → TNF" axis — nice for a review or grant figure. KTP3008 + KTI1020-EN + KTL0100 all port over: anti-ZAR1 rabbit → KTI1020-EN IP → KTL0100 HRP-label for WB, same pipeline as animal T-bet/Foxp3 from KTE9017 piece.
The Bottom Line
Plant total protein extraction stopped being "RIPA + 2% PVPP" the moment your rice BZR1 WB showed 28% CV from phenolic oxidation and chlorophyll-quenched ECL — and "plant RIPA" patches (more PVPP! more ascorbate! X-114!) turn into a 3-batch optimization loop that burns a Friday and still leaves your Arabidopsis rosette sup brown. The ExKine™ Pro Total Protein Extraction Kit for Plant Tissues (KTP3008) from Abbkine ports the animal-Pro detergent backbone (1% NP-40 + 0.3% DOC + 0.5% CHAPS + 0.05% SDS, EDTA-free, 0.1% BSA carrier) into a plant-ready format: 2% PVPP + 1% PVP-40 + 20 mM ascorbate + 5 mM DTT pre-titrated, optional X-114 phase-split for shoot-rich tissues, recovery >90% across rice/Arab/tobacco/maize, BCA CV <7%, and downstream-compatible with KTI1020-EN anti-rabbit IP (BZR1–BIN2, SnRK2–PYR1, ZAR1–RKS1 NLR complexes), KTL0100 HRP labeling (your own anti-BZR1/anti-SnRK2 for zero-ghost WB), and PRM/DIA (pigment-stripped, trypsin-efficient). Whether you're chasing BR-signaling BZR1 dephosphorylation in 24-epiBL-treated rice, drought SnRK2.6 activation in Arab rosette, or NLR–RLCK complexes in N. benthamiana PTI/ETI, it's the plant Pro that doesn't make you re-grind the same 100 mg seedling three Fridays in a row.
Product Reference: KTP3008 – ExKine™ Pro Total Protein Extraction Kit for Plant Tissues
Learn more and order: https://www.abbkine.com/product/exkine-pro-total-protein-extraction-kit-for-plant-tissues-ktp3008/
(For Research Use Only; not for diagnostic procedures in humans.)





