




The Growth Factor That Never Stopped Growing—And the Kit That Measures Only It
Stanley Cohen did not intend to discover epidermal growth factor. He was studying nerve growth factor in mouse submandibular glands in the early 1960s when he noticed that crude gland extracts injected into newborn mice accelerated eyelid opening and tooth eruption. The effect was not neurological. It was epidermal. Cohen isolated the responsible protein, characterized its 53-amino-acid architecture stabilized by three intramolecular disulfide bonds, and demonstrated that it stimulated the proliferation of epidermal and epithelial tissues with a specificity that ruled out non-specific inflammatory mediators. In 1986, he shared the Nobel Prize in Physiology or Medicine with Rita Levi-Montalcini, and the citation explicitly acknowledged that his discovery of EGF had opened an entirely new field of growth factor biology. The molecule that began as an unexpected observation in a mouse colony became the founding member of a family that now includes amphiregulin, epiregulin, betacellulin, heparin-binding EGF, epigen, transforming growth factor-alpha, and the neuregulins—seven ligands, one receptor tyrosine kinase, and a signaling network that controls cell fate decisions in virtually every epithelial tissue in the vertebrate body.
What happened next in the EGF story is a cautionary tale about the distance between a molecule's biological importance and the quality of the tools available to measure it. Human EGF is a 6-kDa secreted peptide generated by proteolytic cleavage from a large transmembrane precursor, pro-EGF, which is expressed most abundantly in the kidney, salivary glands, and gastrointestinal tract. Mature EGF circulates in plasma at low picogram-per-milliliter concentrations, appears in urine at higher levels that correlate with intrarenal EGF mRNA expression and renal functional recovery, and accumulates in tumor microenvironments where autocrine and paracrine EGFR activation drives proliferation, migration, and invasion. A 2024 study published in Scientific Reports confirmed that serum EGF serves as an independent prognostic marker in non-metastatic colorectal cancer, with the circulating concentration distinguishing patient outcomes in a manner that transcript-level measurements could not replicate. Urine EGF has been validated across multiple independent cohorts as a biomarker for chronic kidney disease progression, with the amount of uEGF showing significant correlation with interstitial fibrosis, tubular atrophy, and estimated glomerular filtration rate. In liver disease, serum EGF concentrations correlate with disease severity and have been proposed as an early diagnostic indicator for hepatocellular carcinoma. In depression research, EGF has emerged as a candidate biomarker measurable in cerebrospinal fluid and peripheral blood, with a 2024 review proposing that EGF dysregulation may contribute to the neuroinflammatory component of major depressive disorder.
The analytical challenge that confronts every one of these clinical and translational applications is not that EGF is difficult to detect. It is that EGF is easy to confuse with something else. The EGF family ligands—AREG, EREG, BTC, HB-EGF, TGFA, NRG1-4—share an epidermal growth factor-like domain characterized by a conserved spacing of six cysteine residues that form the three-disulfide-bond scaffold essential for EGFR binding. Amphiregulin and epiregulin, in particular, exhibit sufficient sequence and structural homology with EGF that single-antibody detection formats risk cross-reactivity. A 2014 study in Journal of Biological Chemistry demonstrated that AREG, EREG, and TGFA not only compete with EGF for EGFR binding but induce distinct receptor dimerization kinetics and downstream signaling biases—AREG drives heterodimer formation with ErbB2 through a biphasic kinetic mechanism, while EGF and TGFα bias the receptor toward alternative dimerization partners. When an ELISA quantifies EGF in a biological sample that also contains AREG, which is concurrently upregulated in colorectal cancer and psoriasis, or EREG, which is induced by inflammatory cytokines in epithelial cells, the resulting signal must reflect EGF alone, or the biological conclusion is built on a summed ligand concentration that conflates the outputs of independently regulated genes.
EliKine™ Human EGF ELISA Kit (KTE6005) addresses this specificity challenge through a two-site sandwich architecture that employs two distinct antibodies recognizing separate epitopes on the human EGF molecule. An antibody specific for EGF is pre-coated onto the microplate to capture the analyte from standards and samples. After washing, a biotin-conjugated detection antibody that binds a different EGF epitope is added, followed by proprietary EliKine™ streptavidin-HRP conjugate. A substrate solution then develops color in direct proportion to the amount of EGF captured in the initial step. The color development is stopped, and the absorbance intensity provides a quantitative readout of the EGF concentration in each well. No significant cross-reactivity or interference between human EGF and structural analogues was observed—a specificity specification that the dual-site recognition architecture earns by requiring both antibodies to engage the target for signal generation. A ligand that shares one epitope but not the other produces no signal.
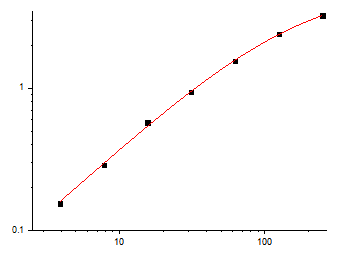
The analytical specifications position the kit within the concentration range that biological research actually demands. The calibration range spans 3.9 pg/mL to 250 pg/mL, with a limit of detection reaching 2 pg/mL. At 2 pg/mL, KTE6005 captures EGF at concentrations that fall below the detection floor of many commercially available ELISA kits, which typically cluster their lower limits of quantification between 7.8 and 31.2 pg/mL. This sensitivity margin enables quantification of EGF in dilute biological matrices—cerebrospinal fluid, bronchoalveolar lavage fluid, conditioned medium from primary cell cultures that secrete EGF at rates far lower than established cell lines—without concentration or pooling strategies that introduce systematic error. The upper boundary of 250 pg/mL accommodates the full physiological range of EGF in plasma and the elevated concentrations observed in tumor-conditioned medium and concentrated urine samples, meaning a single kit serves the researcher who needs to profile EGF across the entire spectrum from healthy baseline to pathological overexpression.
Sample compatibility spans serum, plasma, cell culture supernatants, and other biological fluids. This breadth matters because EGF's biologically relevant distribution is compartmentalized. Plasma EGF reflects systemic production, with concentrations influenced by renal clearance, hepatic metabolism, and the secretory activity of circulating cell populations. Serum EGF captures the platelet-released fraction in addition to the free circulating pool, and the difference between serum and plasma measurements provides information about platelet EGF content that is itself a variable of interest in wound healing research. Cell culture supernatant EGF reveals the secretory behavior of specific cell types—keratinocytes, renal tubular epithelial cells, hepatocytes, tumor cell lines—under defined experimental conditions. Urine EGF reflects intrarenal production and has been validated as a biomarker of functional renal mass and recovery potential. Cerebrospinal fluid EGF reflects central nervous system production and has been implicated in neurogenesis, synaptic plasticity, and the pathophysiology of depression. A kit that accepts all of these sample types serves the nephrologist, the oncologist, the neuroscientist, the wound healing biologist, and the basic scientist within a single standardized workflow.
The component list reflects a complete sandwich ELISA system that ships ready for same-day deployment: Human EGF microplate, Human EGF standard, Human EGF detect antibody, Avidin-HRP, standard diluent, assay buffer, HRP substrate, stop solution, wash buffer, and plate covers. The unopened kit stores at 2–8°C, requiring no -20°C freezer space and subjecting the capture antibody to none of the freeze-thaw cycling that progressively degrades binding affinity in kits stored under suboptimal conditions. Shipping occurs on gel packs with blue ice. The assay follows a multiple-step standard sandwich ELISA protocol with a working time of 3–5 hours, a duration that positions it as a same-day experiment for a technician who begins the first incubation in the morning and reads the plate after lunch.
The protocol notes reflect the operational discipline accumulated through the manufacture and testing of many sandwich ELISA kits. All reagents must warm to room temperature for at least 30 minutes before opening—thermal equilibration that prevents condensation on cold reagent vials from altering concentration. Pipette tips must be pre-rinsed and replaced between samples, standards, and reagents to prevent carryover contamination that is the single largest source of inter-well variability in ELISA workflows. Thorough mixing every 10 minutes using a low-frequency oscillator or gentle hand shaking is specified as critical for result quality. All standards and samples should be assayed in duplicate or triplicate. Do not mix components from different kit lots or use reagents beyond the kit expiration date. Unused wells must be kept desiccated at 4°C in the sealed bag provided. These are the standard operating procedures that separate a standard curve with an R² above 0.99 from a dataset a reviewer will question.
For the oncologist quantifying EGF in preoperative and postoperative serum samples to evaluate its independent prognostic value, the nephrologist measuring urine EGF to predict renal functional recovery in glomerulonephritis patients, the neuroscientist profiling EGF in cerebrospinal fluid from patients with major depressive disorder, the cell biologist measuring EGF secretion from stimulated keratinocyte cultures in a wound healing model, the cancer researcher quantifying autocrine EGF production in a colorectal cancer cell line panel, or the basic scientist performing any experiment in which EGFR pathway activation must be distinguished from non-EGF ligand contributions and intracellular signaling crosstalk, direct EGF quantification is the measurement that converts a pathway-level observation into a ligand-specific mechanism. KTE6005 enables that conversion in a sandwich ELISA format whose sensitivity, specificity, and sample compatibility are documented and whose protocol is transparent enough for a technician to execute on the first attempt.
What distinguishes this kit from the crowded field of EGF ELISA products is not a single specification but the combination of dual-site sandwich specificity in a context where EGF family ligand cross-reactivity is analytically endemic, detection sensitivity that reaches concentrations at which biological differences between health and early disease first become apparent, and sample compatibility that spans the biological matrices in which EGF is actually measured rather than the matrices most convenient to validate. The growth factor that launched the field of growth factor biology can now be quantified without the cross-reactivity that has shadowed its measurement for decades. Cohen isolated EGF from mouse salivary glands in 1962 and opened a field. KTE6005 isolates EGF from the EGF family and delivers a number.
Explore specifications, access the protocol, and place your order here: https://www.abbkine.com/product/elikine-human-egf-elisa-kit-kte6005/





