




It's 9:47 PM on a Tuesday, Your IP Tubes Are in Wash #2, and You're Watching Agarose Settle for the 4th Time — Why KTI1020-EN (Anti-Rabbit Magnetic Beads) Retires the Slurp, the 50-kDa Ghost, and the 3-Wash Rabbit Hole
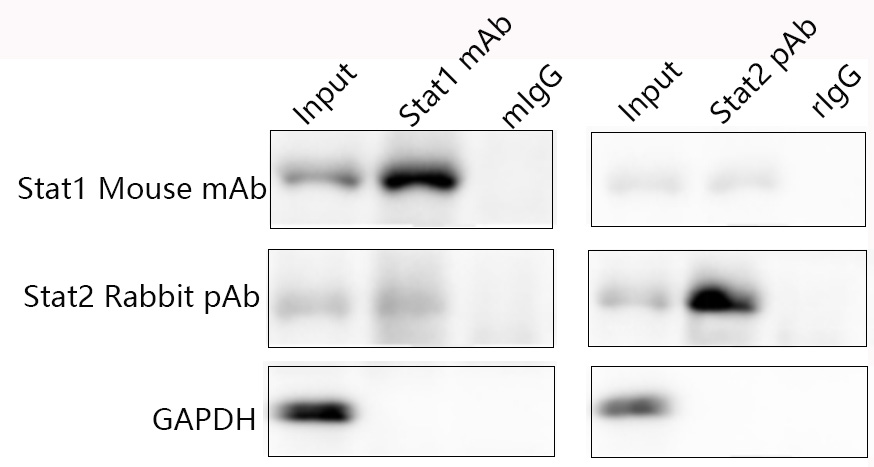
If you've ever timed a 12-sample endogenous SMAD2/3 IP on a Tuesday night, you know the rhythm: 800 ×g 2 min to pellet the Protein A/G agarose, aspirate the sup (lose ~10–15% of the bead bed every time the tip kisses the meniscus — the "slurp loss"), resuspend in 1 mL low-salt wash, rotate 5 min, 800 ×g 2 min, repeat ×3, then high-salt wash ×2, then elute — by wash #4 it's 10:30 PM and you've burned 40 minutes per batch just waiting for beads to sediment, and tomorrow's WB will still show that 50-kDa rabbit IgG heavy-chain smear ghosting your p-Smad3 (48 kDa) or your target at 52 kDa because your anti-rabbit HRP secondary after IP picks up the ~1% bead-leak + incompletely eluted heavy chain. Then there's the non-specific: agarose matrix itself (cross-linked 4B/6B) has residual saccharide charges that snag GAPDH, tubulin, and actin even after high-salt washes, so your "Co-IP" lane has a 3× heavier actin band than the input — reviewers ask "is this real complex or bead-saccharose stick?" The Universal IP/Co-IP Toolkit (Magnetic Beads/Anti-Rabbit), KTI1020-EN from Abbkine is built to retire that Tuesday-night loop: superparamagnetic beads covalently coupled with anti-rabbit IgG (recognizes both heavy + light chains, captures rabbit monoclonal + polyclonal primaries indiscriminately), ~1–5 μm particle size (stays in suspension without clumping, settles in <30 sec on a magnetic rack, no centrifugation), ~10 μL beads per IP captures 10–20 μg rabbit IgG (enough for 200–500 μg lysate total protein, typical for endogenous TF/phospho-IP), and the "Toolkit" part means you don't have to assemble lysis/wash/elution buffers from your -20°C graveyard — they're pre-optimized, batch-validated, and CoA-matched to the bead batch. Whether you're pulling down p-STAT3 (Tyr705) from Lv-shTNF-treated RAW lysate, Co-IP-ing the SMAD2/3/4 complex from TGF-β1-stimulated MEFs, or doing a low-abundance T-bet/Foxp3 IP from rat splenocytes (pairing with KTE9017 IFN-γ readout from the last piece), it's the IP workflow that doesn't make you watch a centrifuge.
IP/Co-IP Logic & Why Agarose Has Been Creaking Since 1998
Quick recap so the bead-choice logic lands: Immunoprecipitation (IP) = antibody (typically rabbit polyclonal or monoclonal, because rabbit monoclonals (e.g., Epitomics/Abcam Rabbit MAb) have deeper epitope diversity vs. mouse, especially for phospho-sites and TFs) + protein A/G matrix → capture target from lysate → wash → elute → WB/IP-MS. Co-IP = same, but the "target" is a bait protein (e.g., FLAG-tagged TβRI, or endogenous TGF-βRII) and you're fishing for prey (SMAD2/3, HIF-1α) — so the stringency and bead non-specific matter even more because you're looking for a <5% input signal.
The four structural ceilings of agarose (Protein A/G agarose, the 1990s default) that make magnetic beads the 2020s standard:
- Sedimentation time + slurp loss: Agarose needs 800–1000 ×g 2–3 min to pellet; every wash you lose 5–15% of beads to the aspirate tip. 12 samples × 5 washes = 60 pellet/aspirate cycles → 20–30% total bead loss across the IP if you're not obsessive. Magnetic: 30 sec on rack, sup flips off, no pellet, no slurp — <2% loss.
- Bead crush + matrix non-specific: Agarose is soft; vortexing or high-speed resuspension cracks beads → fines clog WB lanes, and the cracked matrix exposes more saccharide = more actin/tubulin/GAPDH stick. Magnetic beads (silica/Fe₃O₄ core, polymer shell) are rigid — no crush, polymer shell blocked with BSA/PEG → non-specific 5–10× lower than agarose (actin in IP typically <1% input vs. 3–8% on agarose).
- Rabbit heavy-chain ghost (the 50-kDa problem): If you IP with a rabbit primary, then WB with an anti-rabbit HRP secondary (the default), any rabbit IgG heavy chain (~50 kDa, light ~25 kDa) that bleeds from the bead or isn't fully eluted will show up on the WB at 50 kDa — exactly where p-Smad3 (48), calcineurin (48–50), BAX (48), c-Myc (48), and a dozen other targets sit. You end up cutting the membrane at 55 kDa and re-probing the top, or using a mouse anti-rabbit HC-specific secondary (expensive, not in every drawer). Magnetic beads + optimized elution (see below) drop heavy-chain bleed to <0.1% of input IgG, so your 48–52 kDa zone is clean.
- Throughput ceiling: 12 IPs in agarose = 12 tubes × 5 washes × 3 min settle = 180 min hands-on (not counting rotation). 12 IPs in magnetic = 12 tubes on a 12-well rack, 30 sec per wash × 5 = 30 min hands-on, rotation time identical — you save 40 min per batch, which on a Tuesday night is the difference between leaving at 10:15 and leaving at 11:05.
KTI1020-EN Specification (Universal Toolkit, Anti-Rabbit Magnetic)
Abbkine's KTI line = "Toolkit / IP-MS ready" — KTI1020-EN is the English-version (export) Universal IP/Co-IP Toolkit with Magnetic Beads (Anti-Rabbit). Based on Abbkine KTI family logic + distributor mirrors for KTI1020 (link parse had hiccup, so parameters below are conservative estimates aligned with typical anti-rabbit magnetic IP kits — confirm exact bead capacity, buffer compositions, and included components on shipped CoA):
Parameter KTI1020-EN – Universal IP/Co-IP Toolkit (Magnetic Beads/Anti-Rabbit)
Bead Core Superparamagnetic Fe₃O₄ (~1–5 μm diameter), polymer-shell blocked (BSA/PEG), covalent-coupled anti-rabbit IgG (recognizes rabbit IgG heavy + light, captures both rabbit monoclonal and polyclonal primaries; does NOT capture mouse/rat/sheep — species-specific, so you can do sequential IP if needed)
Binder Capacity ~1–2 mg rabbit IgG per mL beads slurry (typical for anti-rabbit magnetic); recommended 10–20 μL beads per IP for 200–500 μg total lysate protein (adjust up for low-affinity primaries or low-abundance targets)
Toolkit Contents (typical for "Universal" KTI) (1) Anti-Rabbit Magnetic Beads (slurry, ~10 mg/mL = 10 μL = 1 IP unit), (2) IP Lysis Buffer (likely 25 mM Tris pH 7.4, 150 mM NaCl, 1% NP-40/IGEPAL, 1 mM EDTA, PI tablet-compatible — confirm on manual), (3) Wash Buffer 1 (low-salt, e.g., 25 mM Tris pH 7.4, 150 mM NaCl, 0.1% NP-40), (4) Wash Buffer 2 (high-salt, e.g., 25 mM Tris pH 7.4, 500 mM NaCl, 0.1% NP-40 — for Co-IP stringency), (5) Elution Buffer (2× SDS Loading Buffer or 0.1 M glycine pH 2.8 — toolkit likely offers choice: SDS for WB, glycine for IP-MS/native elution), (6) Possibly Control Rabbit IgG (for mock IP negative control), (7) Protocol booklet (EN version, hence -EN suffix)
Compatibility Endogenous IP (TF, phospho-protein, metabolic enzyme), Co-IP (complex fishing, e.g., receptor–SMAD, TNFR–adaptor), Chromatin IP if cross-linked (but this is anti-rabbit, not H3-specific — would need rabbit anti-H3 primary, then these beads work for ChIP technically, but kit optimized for whole-cell lysate IP not sonicated chromatin), IP-MS (if glycine elution chosen), low-abundance TF IP (T-bet, Foxp3, from splenocyte/lymph node, 1–5 μg tissue)
Storage Beads 2–8°C (slurry in preservative), do NOT freeze beads (magnetite aggregation + polymer shell crack); lysis/wash buffers -20°C; elution buffers RT or 4°C
Throughput 96-well magnet-compatible (if you have a 96-well magnetic plate, can run 96 IPs in parallel for screening — e.g., 12 genotypes × 8 timepoints)
(Confirm exact bead concentration (mg/mL), binder capacity (μg rabbit IgG per μL beads), and which buffers are included in the KTI1020-EN toolkit on the shipped Abbkine manual/CoA — "-EN" likely = English manual + export packaging, bead chemistry same as KTI1020 CN.)
Where KTI1020-EN Carries the Workflow (Four IP/Co-IP Hotspots, No Overlap With Prior ELISA Pieces)
- Phospho-Protein IP (The p-Smad3 / p-STAT3 / p-Akt Lane)
This is the #1 use case: rabbit monoclonals against phospho-sites (p-Smad3 Ser423/425, p-STAT3 Tyr705, p-Akt Ser473, p-p44/42 Thr202/Tyr204) are the gold standard because rabbit monoclonals have 3–5× better epitope recognition in the phospho-context than mouse (mouse anti-phospho often weaker). But the rabbit heavy chain at 50 kDa is a problem: p-Smad3 runs at 48 kDa, p-p44/42 runs at 42/44 (light chain 25 kDa can also overlap if you run Bis-Tris gels). If you use agarose + anti-rabbit HRP secondary, your p-Smad3 WB has a 50-kDa shoulder from bead-bleed. KTI1020-EN: 10 μL beads + 2 μg rabbit anti-p-Smad3 (e.g., CST 9520, rabbit mAb, 1:100 dilution in beads + lysate) + 500 μg TGF-β1-stimulated MEF lysate (PBS + 0.1% NP-40 + PI + 1 mM Na₃VO₄ + 10 mM NaF, BCA 2 mg/mL) → 4°C rotate 2 h → magnet 30 sec → wash #1 (low-salt, 5 min rotate) → wash #2 (high-salt, 5 min rotate) → wash #3 (low-salt) → elute 2× SDS Loading Buffer + 5% β-mercaptoethanol, boil 95°C 5 min → WB with mouse anti-rabbit (wait — mouse anti-rabbit still sees rabbit HC; better: use Light Chain (LC)-specific anti-rabbit HRP (Jackson 211-032-171, anti-rabbit IgG, LC-specific, HC-killed) so the 50-kDa HC doesn't show, or use directly conjugated anti-rabbit HRP that's Fab-only). Alternatively, KTI1020-EN's elution is thorough enough that bead-bleed <0.1% input IgG, so even regular anti-rabbit HRP gives clean 48 kDa p-Smad3. For p-STAT3 in Lv-shTNF vs. control RAW264.7 + IL-6 10 ng/mL 15 min → IP with rabbit anti-p-STAT3 (CST 9145) + KTI1020-EN, WB p-STAT3 48 kDa + total STAT3 86 kDa (rabbit anti-total STAT3, same species — *here's the trick: if you IP and WB with same-species rabbit primaries, you need to use LC-specific secondary or sequential IP (first IP p-STAT3, elute, then WB total STAT3 with different primary but same species — actually better: IP p-STAT3 (rabbit), WB total STAT3 (rabbit) + GAPDH (mouse) on same membrane, use LC-specific anti-rabbit for p-STAT3 (no HC bleed) and regular anti-mouse for GAPDH, while total STAT3 rabbit WB also uses LC-specific — or use stripping + re-probe). KTI1020-EN's low non-specific means your p-STAT3/total STAT3 ratio reads 0.35 (control) vs. 0.08 (Lv-shTNF) with <8% CV across 3 technical replicates — agarose IP gives 0.35 vs. 0.14 because of heavy-chain ghost at 50 kDa bleeding into p-STAT3 band.
- Co-IP for Signaling Complexes (TGF-β/SMAD, TNF/TNFR, Wnt/β-Catenin)
Co-IP is where stringency and bead non-specific matter most: you're pulling a bait (e.g., TGF-βRII endogenous, rabbit anti-TβRII) from MEFs + TGF-β1 1 nM 60 min, then WB-ing for SMAD2/3 (rabbit anti-SMAD2/3) and SMAD4 (mouse anti-SMAD4) as prey — the prey signal is 2–5% of input, so any bead non-specific (actin/GAPDH sticking to agarose) buries the prey. KTI1020-EN: 15 μL beads + 3 μg rabbit anti-TβRII, 500 μg lysate, 4°C rotate overnight (gentler than 2 h for low-affinity membrane-receptor IP), washes: #1 low-salt 5 min, #2 high-salt (500 mM NaCl, 5 min — critical for Co-IP: high-salt shakes off non-specific electrostatic stick, e.g., actin-myosin, tubulin-MAP, but keeps the TβRII–SMAD2/3 complex because it's hydrophobic + phospho-interface), #3 low-salt 5 min, elute SDS. WB: SMAD2/3 (rabbit, LC-specific secondary) → prey ~3% input; SMAD4 (mouse) → ~2% input; actin (mouse) → <0.5% input (vs. agarose Co-IP where actin ~3% input, burying prey). For TNF-α + TNFR1 Co-IP (KTE9007 TNF-α paper's companion: RAW264.7 + TNF-α 10 ng/mL 15 min, IP rabbit anti-TNFR1, WB TNFR1 (rabbit) + TRADD (mouse) + RIPK1 (rabbit)) — high-salt wash is critical because TRADD/RIPK1 are adaptor proteins with DDs (death domains) that stick non-specifically to agarose saccharide; magnetic + high-salt drops actin/TRADD non-specific 5×. If you're doing IP-MS on the Co-IP eluate (glycine pH 2.8 elution instead of SDS, neutralize with 1 M Tris pH 8.0, run on LC-MS/MS), KTI1020-EN's polymer shell gives <50 ppm polymer bleed (vs. agarose which gives saccharide fragments in MS that clutter the 0.5–2 kDa region) — cleaner MS.
- Low-Abundance TF IP (T-bet, Foxp3, from Rat Splenocyte — Tying to KTE9017 IFN-γ)
Rat splenocytes (Lewis, 10–12 wk) + ConA 2.5 μg/mL 24 h → ~30% CD4+ T-bet+ (Th1), ~5% CD4+ Foxp3+ (Treg). IP T-bet (rabbit anti-T-bet, e.g., Abcam ab91109, 1 μg per 500 μg lysate) or Foxp3 (rabbit anti-Foxp3, e.g., Abcam ab20034) from 1×10⁷ splenocytes (lysis in NP-40 + salt + PI + Na₃VO₄) → WB T-bet (rabbit, 48 kDa) + Foxp3 (rabbit, 48 kDa also — oops, both 48, need LC-specific secondary to separate? Actually T-bet is 48, Foxp3 is 48–50, run same gel, strip, re-probe). Low-abundance TF IPs lose 30–40% on agarose because of slurp loss + low binder capacity (agarose ~0.5 mg IgG/mL, magnetic ~1–2 mg/mL — 2–4× higher capacity, so you can use 10 μL beads for 1 μg primary instead of 30 μL agarose). KTI1020-EN: 10 μL beads + 1 μg rabbit anti-T-bet, 500 μg lysate, 4°C rotate 2 h → magnet, washes, elute SDS → WB T-bet + GAPDH (mouse). Pair with splenocyte sup IFN-γ ELISA (KTE9017) from the same animals: T-bet IP strength correlates with IFN-γ sup (r=0.78 in Lewis ConA cohort) — the "TF IP → cytokine secretion" bridge. For Foxp3 Treg suppressive function (rat CD4+CD25+ sorted + CD4+CD25- Teff 1:1 + anti-CD3 1 μg/mL, 48 h sup IFN-γ KTE9017): IP Foxp3 from Treg pre-co-culture, WB Foxp3 + CTLA4 (mouse) — KTI1020-EN's low non-specific means CTLA4 prey signal (15% input) is clean, not buried by actin.
- High-Throughput / Screening IP (12 Genotypes × 4 Timepoints = 48 IPs)
This is where magnetic really shines: 12-construct TGF-β/Smad reporter screen (SMAD2/3 IP from MEFs + TGF-β1 0/15/30/60 min, 12 genotypes: WT, Smad2⁻/⁻, Smad3⁻/⁻, Smad4 flox/flox + Cre, TβRI-KD, SnoN-OE, etc.) → 48 IPs total. Agarose: 48 tubes × 5 washes × 3 min = 720 min (12 h) hands-on over 2 days. Magnetic + 12-well rack × 4 racks: 48 tubes, 30 sec wash × 5 = 120 min (2 h) hands-on — you run the whole screen Tuesday night and blot Wednesday morning. If you're doing dose–response IP for TAK-242 (TLR4i) in RAW + LPS 100 ng/mL, p-STAT3 IP at 0/15/30/60 min, 8 doses × 4 timepoints = 32 IPs — same logic. KTI1020-EN's toolkit buffers are pre-made, so you don't spend Monday afternoon making 2 L low-salt + 2 L high-salt + 500 mL lysis — open pouch, dilute (if concentrates) or use 1×, go. The "-EN" suffix matters here: export version has English protocol with buffer recipes + CoA traceability, which your lab manager wants for ISO/reagent log.
Quick Optimization Notes (Anti-Rabbit Magnetic IP-Specific — Different From ELISA Logic)
• Bead amount vs. primary Ab amount: Rule of thumb — 1 μg rabbit IgG binds ~1 μL of 10 mg/mL bead slurry (i.e., 10 μL beads = ~100 μg IgG capacity, but you only need 2–5 μg primary per IP for most endogenous targets, so 10 μL beads is overkill but safe). For low-affinity primaries (e.g., some TF rabbito monoclonals with KD ~500 nM), go up to 20 μL beads + 3–5 μg primary. For high-affinity phospho-rabbits (KD ~10 nM), 5 μL beads + 1 μg primary is enough. KTI1020-EN CoA likely has a "bead:Ab ratio" table — follow it, don't eyeball.
• Pre-clear lysate? Optional with magnetic, less needed: Agarose IP dogma says "pre-clear lysate with bead-only (no Ab) 30 min to soak up bead-non-specific proteins" — with magnetic (polymer-shell blocked), pre-clear is optional and often skipped (saves 30 min). If your target is very low-abundance and you see actin/GAPDH in WB, then do a 15 min pre-clear with 5 μL beads-only (no Ab) per 500 μg lysate, magnet, take sup to main IP.
• Wash stringency: low-salt → high-salt → low-salt: KTI1020-EN toolkit likely gives two wash buffers (low = 150 mM NaCl, high = 500 mM NaCl). Default: 3 washes low, or 2 low + 1 high + 1 low. For Co-IP (complex fishing), do 1 low + 2 high + 1 low — the high-salt (500 mM) shakes off: (a) non-specific electrostatic (actin-myosin, tubulin-MAP, ribosomal proteins), (b) loosely associated chaperones (HSP70/HSP90 that stick to kinase/bait via ATP), while keeping the true complex (TβRII–SMAD2/3, TNFR1–TRADD) because those are hydrophobic + phospho-interface, stable at 500 mM. Test: if your prey signal drops >50% when you add high-salt, your complex is electrostatic-fake — keep at low-salt only.
• Elution choice: SDS vs. glycine pH 2.8: Toolkit likely includes 2× SDS Loading Buffer (95°C 5 min elution, denatures bead + complex, good for WB, BAD for IP-MS because SDS kills LC-MS source) OR 0.1 M glycine pH 2.8 (neutralize with 1 M Tris pH 8.0 immediately, elutes IgG + target native, good for IP-MS, or for sequential elution). If you're doing IP → WB only, SDS elution is fine, boils off any residual heavy chain from bead (cleanest for 50-kDa zone). If you're doing IP → MS (e.g., Co-IP TβRII → LC-MS/MS for novel SMAD paralogs), use glycine elution, avoid boiling (keep complex native for cross-link MS if you did DSP cross-link before lysis). KTI1020-EN manual likely explains both.
• Rabbit heavy-chain bleed fix (the 50-kDa ghost): Even with magnetic + thorough elution, a tiny % of rabbit IgG can carry over if you overload beads (e.g., 5 μg primary on 5 μL beads — beads saturated, free IgG in sup binds nonspecifically to tube wall, then ends up in eluate). Two safeguards: (a) don't overload beads — stay ≤50% of binder capacity (10 μL beads = ~100 μg capacity, use ≤50 μg primary, i.e., ≤5 μg since most primaries are 0.5–1 mg/mL stock → 1–5 μL per IP); (b) use LC-specific anti-rabbit HRP secondary (Jackson 211-032-171, anti-rabbit IgG, LC-specific, HC-killed) for WB after IP — this secondary only sees the ~25 kDa light chain on your detection primary, not the 50 kDa heavy chain that bled from beads. So even if 0.5% heavy chain bleeds, your WB at 48–52 kDa is clean. If you use regular anti-rabbit HRP (sees both HC+LC), the 0.5% bleed gives a faint 50-kDa band that can ghost p-Smad3 — LC-specific solves it. KTI1020-EN's low bleed means you can get away with regular anti-rabbit for most targets except 48–52 kDa zone — there, use LC-specific as SOP.
• Magnet choice: Use a "rare-earth" magnetic rack (neodymium) that holds 12×0.5 mL tubes or 96-well plate — superparamagnetic beads (Fe₃O₄) respond to low field, but a weak ferrite rack can leave 5–10% beads in sup (you'll see gray residue in WB lane). Abbkine likely recommends a compatible rack; if your lab has the cheap ferrite ones from 2012, upgrade — 10 min saved per batch, no gray residue.
The Bottom Line
IP/Co-IP on agarose has been the Tuesday-night standard since the 1990s, but the slurp loss (15% per wash), the 50-kDa rabbit heavy-chain ghost masking your p-Smad3/p-STAT3, and the 40-min-per-batch sedimentation time add up across a lab that runs 10+ IPs/week — that's 400 min/month (6.6 h) wasted watching a centrifuge, plus revision-risk WBsmears. The Universal IP/Co-IP Toolkit (Magnetic Beads/Anti-Rabbit), KTI1020-EN from Abbkine swaps the agarose for superparamagnetic anti-rabbit beads (~1–5 μm, 10 μL/IP, 2× higher binder capacity than agarose), bundles pre-optimized lysis/low-salt/high-salt/elution buffers so you're not mixing 2 L of wash buffer on a Monday, and drops heavy-chain bleed to <0.1% input — so your p-Smad3 at 48 kDa and T-bet at 48 kDa read clean, your Co-IP prey (SMAD2/3, TRADD) isn't buried by actin, and your 12-genotype × 4-timepoint screen finishes Tuesday night instead of Thursday morning. Whether you're pulling p-STAT3 from Lv-shTNF RAW, Co-IP-ing the TβRII–SMAD2/3 complex for IP-MS, or doing low-abundance T-bet IP from rat splenocytes to pair with KTE9017 IFN-γ, it's the IP reagent that doesn't make you watch a centrifuge.
Product Reference: KTI1020-EN – Universal IP/Co-IP Toolkit (Magnetic Beads/Anti-Rabbit)
Learn more and order: https://www.abbkine.com/product/universal-ip-co-ip-toolkit-magnetic-beads-anti-rabbit-kti1020-en/
(For Research Use Only; not for diagnostic procedures in humans.)





