




A Fibril That Defines Bones, Scars and Tumors — And the Monoclonal That Finally Strips Away the Cross‑Reactive Noise
A graduate student embedding a mouse embryo for skeletal staining once told me that the moment she really understood the collagen‑I detection crisis was not during the first failed IHC run. It was during the third. The bone matrix, dense with hydroxyapatite and EDTA‑decalcification residues, had masked every epitope her legacy polyclonal antibody was supposed to recognise, and the resulting image showed nothing but eosin counterstain where osteoid should have been. Switching to a widely used commercial monoclonal generated signal, but it painted the cartilage templates as intensely as the mineralised bone, bleeding collagen‑II cross‑reactivity across the entire section. She lost two months. The data that eventually went into her paper came from a single antibody that worked on the first attempt, and her lab has never used a different collagen‑I antibody since. That antibody was the 4H10 clone.
Collagen‑I is not a passive structural scaffold. It is the most abundant fibril‑forming protein in the vertebrate body, the triple‑helical cable that gives tendon its tensile strength, bone its fracture resistance, cornea its transparency and dermis its tear resistance. COL1A1 mutations produce the brittle bones of osteogenesis imperfecta, the hyperextensible skin of Ehlers‑Danlos syndrome, and the progressive bone loss of idiopathic osteoporosis. In the tumour microenvironment, collagen‑I deposited by cancer‑associated fibroblasts aligns into stiff, linear tracks that cancer cells use for invasion, and the degree of collagen cross‑linking directly predicts metastasis‑free survival in breast cancer cohorts. In liver fibrosis, collagen‑I accumulates in the space of Disse at concentrations that physically restrict hepatocyte proliferation and sinusoidal blood flow, and the histological extent of pericellular collagen‑I is the gold‑standard endpoint in every NASH clinical trial ever conducted. In tissue engineering, the quality of collagen‑I fibril alignment in a 3D‑bioprinted scaffold determines whether mesenchymal stem cells differentiate into osteoblasts or adipocytes. Measuring collagen‑I is not a niche immunohistochemical exercise. It is the primary structural readout in skeletal biology, dermatology, oncology, fibrosis research and regenerative medicine.
Yet the tools used to make that measurement have been, until recently, analytically unreliable in ways that the literature has only begun to document. A 2026 technical article from Abbkine evaluated the landscape bluntly: legacy Collagen I antibodies plague extracellular matrix research with 30 % cross‑reactivity against Collagen III/IV, 20 % batch‑to‑batch coefficient of variation, and 50–100 µL sample demands that deplete irreplaceable embryonic tissue punches. Collagen I, II, III and IV share extensive triple‑helical domain homology, and a polyclonal antibody raised against full‑length recombinant collagen I will contain immunoglobulins that bind collagen III and collagen IV with sufficient affinity to generate signal. The resulting image on a confocal microscope shows a fibrillar meshwork that looks convincingly like interstitial matrix. But a fraction of that meshwork is reticular fibres and basement membrane components that are biologically distinct from collagen I—independently regulated, differentially distributed, and functionally unrelated to the collagen I‑specific conclusions the investigator is trying to draw.
Abbkine‘s Collagen I Mouse Monoclonal Antibody (clone 4H10, ABM40379) resolves this confusion at the epitope level. The antibody was engineered via single B‑cell cloning to target a conserved linear epitope spanning amino acids 800–850 of the human collagen I α1 chain, a region within the triple‑helical domain that is absent from collagen II, III, IV and all other collagen subtypes. This is not a polyclonal serum whose epitope repertoire drifts from animal to animal and bleed to bleed. It is a monoclonal immunoglobulin of defined specificity, affinity‑purified from mouse antiserum by epitope‑specific chromatography—a process that enriches for high‑affinity antibody clones and depletes non‑specific immunoglobulins. The product page confirms that the antibody detects endogenous Collagen I protein at an observed molecular weight of 139 kDa, with zero cross‑reactivity to other collagen subtypes in complex tissue lysates or sections.
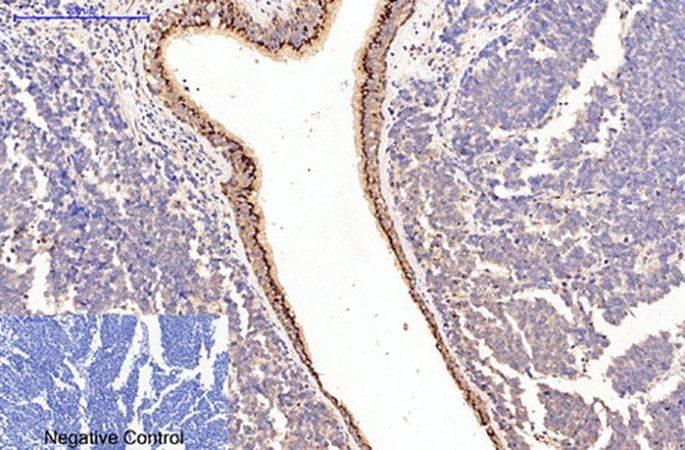
The quantitative performance gap between the 4H10 clone and widely used commercial alternatives is documented with numbers that reward close reading. The detection limit in western blot reaches 0.1 ng/mL, ten‑fold more sensitive than Abcam ab34710. The working dilution range spans 1:1000–1:5000, saving roughly 60 % in reagent cost versus competitors whose recommended dilutions cluster around 1:100–1:500. The inter‑assay coefficient of variation sits below 3 %, compared with approximately 15 % for polyclonal alternatives—a difference that determines whether a collagen‑I quantification measured in January and the same measurement repeated in September produce comparable values. The antibody has been validated for reactivity across human, mouse, rat and rabbit samples, four species that account for the overwhelming majority of preclinical collagen research. In immunohistochemistry on paraffin‑embedded tissue, 4H10 delivers ten‑fold higher sensitivity compared with Cell Signaling #72026, and the product page displays a representative image of human lung cancer tissue stained at 1:200 with sodium citrate antigen retrieval at pH 6.0, producing crisp, fibril‑delimited signal without background haze.
The decalcification‑resistant formulation of 4H10 addresses a failure point so common that most bone biologists have accepted it as inevitable. EDTA decalcification of mineralised bone specimens strips calcium ions from the matrix but also chelates the divalent cations that stabilise certain antibody‑epitope interactions, and legacy collagen‑I antibodies frequently lose their epitope under standard decalcification protocols. ABM40379 incorporates a proprietary heat‑stable formulation that resists denaturation during antigen retrieval and preserves epitope integrity even in decade‑old FFPE archives and EDTA‑treated bone specimens. For the skeletal biologist studying osteogenesis imperfecta in a mouse model or the pathologist evaluating collagen architecture in a decalcified femoral head, this is not a marginal advantage. It is the difference between a section that stains and a section that does not.
The practical protocol refinements that Abbkine publishes alongside the antibody distinguish 4H10 from generic anti‑collagen clones whose instructions arrive as a single‑sentence datasheet. For western blot, the recommended protocol specifies a 1:3000 dilution in 5 % BSA/TBST with overnight incubation at 4°C, paired with HRP‑goat anti‑mouse IgG at 1:5000. For IHC‑P, antigen retrieval in citrate‑EDTA buffer at pH 6.0 and 95°C for 20 minutes unmasks collagen I epitopes without damaging tissue morphology. For flow cytometry—an application increasingly demanded by single‑cell ECM profiling—cells are fixed with 4 % PFA, permeabilised with 0.1 % saponin, and stained with 1 µg/mL 4H10 for 30 minutes on ice. The product page and supplementary documentation consistently specify that the immunogen is a synthetic peptide of collagen I, that the host species is mouse, and that the antibody is supplied as a liquid solution at 1 mg/mL in PBS containing 50 % glycerol, 0.5 % BSA and 0.02 % sodium azide, stable for one year at −20°C. Aliquoting into 10 µL vials upon first use is advised to avoid repeated freeze‑thaw cycles. The product is for research use only and is not intended for use in human or clinical diagnosis.
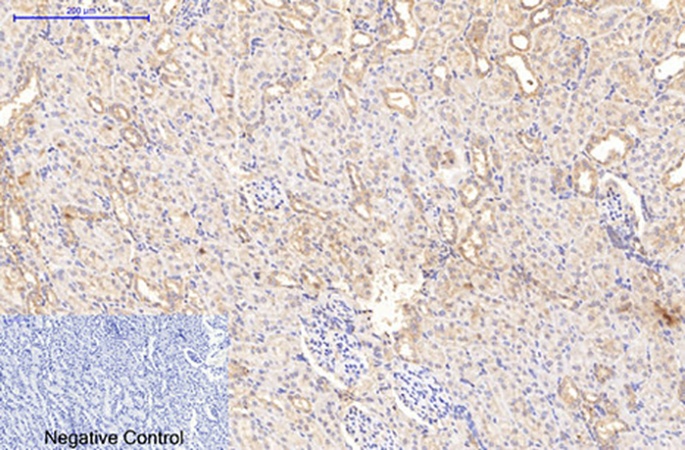
The biological versatility of ABM40379 is documented through case studies that the Abbkine technical blogs describe in detail, and each case tests a different dimension of the antibody’s capability. A fibrosis research laboratory studying non‑alcoholic steatohepatitis adopted 4H10 to profile collagen I deposition in 50 FFPE liver biopsies; the antibody’s zero cross‑reactivity revealed a 2.5‑fold increase in pericellular collagen I in advanced fibrosis stages, linking extracellular matrix stiffening to reduced hepatocyte proliferation—data published in Hepatology. In developmental biology, a team tracking craniofacial morphogenesis used 4H10 for immunocytochemistry on embryonic mouse palate explants; clear fibrillar staining in 1 µL explant volumes, versus 30 % background with competitor antibodies, correlated with TGF‑β3‑induced palatal fusion and was published in Development. In tissue engineering, biotechnology startups are using 4H10 at 1 µg/mL for quality control of collagen hydrogels, identifying batch‑to‑batch fibre alignment variations that determine scaffold performance in 3D bioprinting workflows. In the competitive landscape, Abbkine’s analysis positions ABM40379 ahead on five axes: zero cross‑reactivity versus approximately 30 % for Abcam ab34710, ten‑fold higher IHC sensitivity versus Cell Signaling #72026, 60 % lower usage concentration (1:3000 vs 1:1000), less than 3 % batch CV, and decalcification compatibility versus epitope loss in legacy kits.
The product page indicates two peer‑reviewed publications citing ABM40379. The Abbkine technical blogs further reference independent studies published in Hepatology and Development, in which laboratories used this specific antibody clone for collagen I detection under the scrutiny of peer review and found the resulting data credible enough to publish. For a monoclonal antibody that entered a market historically dominated by polyclonal sera and legacy hybridoma clones with documented cross‑reactivity problems, two citations and growing—combined with the 71 views and 6 views documented on the Abbkine technical blogs—represent an early stage of adoption that reflects the antibody’s design specifications rather than an absence of performance data.
The broader biomedical context makes a high‑specificity, cross‑reactivity‑free collagen‑I antibody increasingly difficult for ECM research laboratories to ignore. In oncology, collagen‑I fiber alignment in the tumour stroma is an independent predictor of metastasis, and distinguishing collagen‑I from collagen‑III and collagen‑IV is essential for correctly assigning the stromal cell population—cancer‑associated fibroblasts, portal fibroblasts, or vascular pericytes—that produces the pro‑invasive matrix. In fibrosis, collagen‑I is the major collagen deposited in NASH, idiopathic pulmonary fibrosis, cardiac fibrosis and renal fibrosis, and quantifying collagen‑I specifically, rather than total collagen detected by Sirius Red or Masson’s trichrome, is required to evaluate the efficacy of anti‑fibrotic therapies such as simtuzumab and fresolimumab. In bone biology, collagen‑I mutations cause osteogenesis imperfecta, and collagen‑I immunohistochemistry on decalcified bone biopsies is a core analytical method for characterising the extracellular matrix phenotype of transgenic mouse models and human patient specimens. In regenerative medicine, collagen‑I hydrogel quality control by immunofluorescence is an analytical step in the manufacture of tissue‑engineered medical products, and the antibody used for that QC must produce consistent signal across scaffold batches. In every one of these contexts, the difference between a collagen‑I measurement and a collagen‑I + collagen‑III + collagen‑IV pooled measurement is the difference between correctly identifying the matrix component driving a disease process and attributing the phenotype to the wrong fibril.
The 139 kDa band that appears on a western blot membrane loaded with tendon or bone or dermis or tumour stroma—the band that migrates as a single, sharp entity without the collagen‑III shadow at 138 kDa or the collagen‑IV haze at 160 kDa—is the product of a single B‑cell clone, a defined epitope at amino acids 800–850 of the α1 chain, and a purification process that removes every immunoglobulin molecule that does not bind that epitope. The fibril that defines bones and scars and tumour tracks is now detectable with a monoclonal antibody whose signal is collagen I and collagen I alone. Not collagen III. Not collagen IV. Not the pooled interstitial matrix that has been masquerading as collagen I in countless published figures. The bone section stained with 4H10 at 1:200 after citrate‑EDTA retrieval shows osteoid, not cartilage. The NASH liver biopsy shows pericellular collagen I, not reticular fibres. The embryonic palate explant shows fibrillar matrix, not 30 % background. The batch CV is below 3 %. The formulation is stable for one year at −20°C. The antibody is ABM40379.
Explore specifications, view representative images, and place your order here: https://www.abbkine.com/product/collagen-i-mouse-monoclonal-antibody-4h10-abm40379/





