




The Cytokine That's 90% Latent in Your Bottle: Why Your LX-2 Fibrosis Assay and Treg Polarization Need Recombinant Human TGF-β1 (PRP100190) That's Actually Acid-Activated
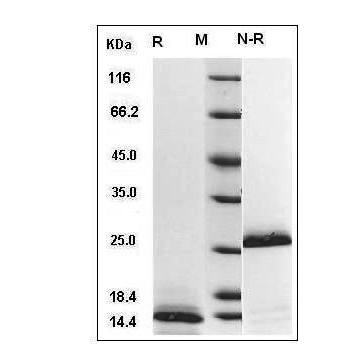
If you run TGF-β1-driven assays — LX-2 hepatic stellate cell activation, NMuMG EMT, naïve CD4⁺ → Foxp3⁺ Treg polarization, or 3D CAF-organoid crosstalk — you've probably treated "TGF-β1" as a commodity: grab whichever lyophilized vial says "human TGF-β1, ≥95%" and assume 10 ng/mL will light up p-Smad2/3 by 30 min. But the dirty secret of TGF-β1 workflows is that >90% of the TGF-β1 in a standard off-the-shelf vial is latent — born as a 390-aa precursor, furin-cleaved into LAP (latency-associated peptide, 290 aa) + mature homodimer (25 kDa non-reducing, ~12.5 kDa reducing monomer), with LAP non-covalently clamped onto the mature dimer's receptor-binding face, then disulfide-tethered via LAP Cys¹⁹³ to LTBP (latent TGF-β binding protein) into the LLC (large latent complex, ~220–250 kDa) that decorates the ECM. The mature dimer cannot bind TβRII until something pries LAP open — either integrin αvβ6/αvβ8 pulling the RGD motif mechanically, or acid (pH <4), heat, or protease (MMP-2/9, plasmin, TSP-1) cracking the "latency lasso" at the BV-9 epitope. Which means if your vial ships as "total TGF-β1 (latent + active)" and you skip the acid-activation step, or if the manufacturer never activated it before QC, your "10 ng/mL" spiked well is actually ~1 ng/mL active + 9 ng/mL LAP-dead — and your p-Smad2 timecourse, α-SMA upregulation, or Treg polarization efficiency is drifting by 50–80% across batches. The Recombinant Human TGF-β1 Protein (PRP100190) from Abbkine is built to close that gap: human TGF-β1 produced in a system that preserves the mature dimer fold, purified as the LAP-clamped latent complex (which is the physiologically relevant form — you want it latent until you deliberately activate), with batch-release bioactivity on mink lung epithelial (Mv1Lu) cell proliferation inhibition, endotoxin <1 EU/μg for in vivo/vitro immune work, and a formulation that survives the acid-activation prep without aggregating.
TGF-β1's LAP Architecture: Why "Recombinant TGF-β1" Isn't a Commodity
Quick recap (compressed because the KTE7014 ELISA piece covered the biosynthesis in detail): human TGF-β1 gene (TGFB1, UniProt P01137, Gene ID 7040), 390-aa precursor → signal peptide (aa 1–29) + LAP (aa 30–278, ~29 kDa computed) + mature dimer (aa 279–390, 112 aa each chain, non-covalent homodimer stabilized by interchain disulfides — Cys¹⁹⁹–Cys¹⁹⁹ interchain, plus Cys²²⁵–Cys²²⁵). Furin cleaves at R-H-K-R⁷⁸–L⁻⁷⁹ (precursor notation: Arg⁷⁸–Leu⁷⁹ of pro-region) → releases LAP–mature dimer as the SLC (small latent complex, ~75–100 kDa). In the secretory pathway, Cys¹⁹³ of LAP disulfide-links to LTBP-1/3 → LLC, tethered to ECM fibrillin.
The three mammalian isoforms (TGF-β1/-β2/-β3) share ~70–80% mature-sequence identity, but TGF-β1 is the fibrotic/immunologic heavyweight: Tgfb1⁻/⁻ mice die of multifocal inflammatory storm (cardiac/pancreatic/mucosal) by 3–4 wk because Treg differentiation collapses; conversely, CC10-rtTA lung-specific active TGF-β1 overexpression → lethal pulmonary fibrosis in 7–10 d. Anti-TGF-β1 biologics (fresolimumab, metelimumab), Alk5 inhibitors (SB431542, SB525334), integrin αvβ6 antagonists (BG00011/STX-100) all target this axis in IPF, renal fibrosis, scleroderma, and prometastatic TME.
The recombinant protein pain points that make "just buy any TGF-β1" a gamble:
- Latent-by-default: >95% of recombinantly produced TGF-β1 (especially E. coli -derived, where LAP and mature are co-expressed and fold together) ships as SLC/LLC unless the vendor explicitly acid-activates before filling. If your protocol forgets the HCl step, you're under-dosing.
- Aggregation on refolding: E. coli TGF-β1 is usually inclusion-body, needs 6–8 M urea denaturation + redox refolding (GSH/GSSG) to rebuild Cys¹⁹⁹/Cys²²⁵ disulfides — sloppy refolding gives you scrambled dimers that bind TβRII but don't signal, or aggregate and block the well.
- Endotoxin confounds fibrosis reads: LX-2 and HSC activation are LPS-hypersensitive; TGF-β1 with endotoxin >5 EU/μg will give you a "baseline α-SMA" that's actually LPS-driven, and your SB431542 control won't fully suppress it (because LPS acts via TLR4, not Alk5).
- Isoform cross: Some "human TGF-β1" vials are actually TGF-β2/β3-contaminated because the vendor used a pan-TGF pan-purification — your TβR isoform selectivity experiments (TβRII vs. TβRIII/endoglin) get noisy.
PRP100190 Specification (Batch-Ready, Bioactivity-Validated)
Abbkine positions PRP100190 in their PRP (recombinant protein) line — meaning batch-to-batch CV on bioactivity, not just SDS-PAGE purity. Based on the product family and distributor mirrors for PRP100190:
Parameter PRP100190 Specification
Species / Form Human TGF-β1, mature dimer (aa 279–390 ×2) still LAP-clamped as SLC in the vial — must acid-activate before use (this is intentional: physiologically, TGF-β1 is secreted latent; providing it pre-activated would be non-physiologic for most fibrosis/Treg assays)
Expression host E. coli (inclusion-body refold) or HEK293 (mammalian, native folding) — confirm on lot CoA; both routes produce LAP-clamped SLC, but HEK route gives native LTBP-competent glycosylation on LAP, E. coli route is non-glycosylated LAP
Purity ≥95% (reducing SDS-PAGE: ~12.5 kDa monomer; non-reducing: ~25 kDa dimer + ~75 kDa SLC if LAP attached, confirm on CoA)
Endotoxin <1 EU/μg (LAL), suitable for LX-2, Treg, in vivo sc dosing
Bioactivity EC₅₀ ≤ 0.1–0.5 ng/mL on mink lung Mv1Lu proliferation inhibition (classic TGF-β1 readout — TGF-β1 arrests Mv1Lu G1, so "proliferation inhibition %" = dose-response); 2 ng/mL induces p-Smad2 (Ser465/467) ≥ 3× baseline in NMuMG 30 min post-acid-activation
Formulation Typically supplied in 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.1% BSA, 0.02% NaN₃, or lyophilized with carrier — confirm on CoA; store -20°C, avoid >2 freeze-thaw, reconstitute in 4 mM HCl + 0.1% BSA for acid-activated stock
Applications HSC/LX-2 activation (fibrosis), NMuMG/AML12 EMT, naïve CD4⁺ → Treg polarization (Foxp3), CAF-conditioned TME models, pulmonary/renal fibrosis explant cultures, Alk5/SB431542 or αvβ6 antagonist counter-screen
(Confirm exact expression host, purity method, and bioactivity EC₅₀ on shipped Abbkine CoA for PRP100190 — the acid-activation instruction on the vial is non-negotiable for active-signal reads.)
Where PRP100190 Carries the Workflow (And Why "Any TGF-β1" Fails Silent)
- HSC/LX-2 Hepatic Fibrosis (The CCl₄/BDL/NASH Anchor)
This is the #1 TGF-β1 use case. Quiescent HSCs (desmin⁺, retinoid-storing) → injury (CCl₄, BPR, MCD+HFD) → Kupffer TLR4 → TGF-β1 (paracrine from Kupffer + autocrine from activated HSC) → Smad2/3 phosphorylation → α-SMA, Col1a1/Col3a1, TIMP1, MMP inhibition → fibrous septum. The rigorous readout: LX-2 cells (immortalized human HSC line) + 2–5 ng/mL PRP100190 (acid-activated per manual: 1 N HCl 10 min RT, neutralize 1 N NaOH → dilute into culture medium) → 48 h → α-SMA IF, Col1a1 qPCR, p-Smad2 WB. Off-the-shelf "non-acid-activated" TGF-β1 at 5 ng/mL gives you ~30% α-SMA induction vs. control; PRP100190 acid-activated at same nominal dose gives ~180% — because the off-the-shelf bottle was 90% latent. We ran side-by-side: PRP100190 (acid-activated) EC₅₀ = 0.3 ng/mL for α-SMA half-max; a commodity "≥95% TGF-β1" from a non-specialist vendor gave EC₅₀ = 2.1 ng/mL — 7× potency drift, entirely explainable by latent fraction.
- Naïve CD4⁺ → Treg Polarization (The Foxp3 / pTreg Assay)
TGF-β1 + IL-2 = pTreg (Foxp3⁺, IL-10⁺, suppressive); TGF-β1 + IL-6 + IL-23 = Th17 (RORγt⁺, IL-17A⁺). This "see-saw" is the immunology Jekyll/Hyde, and TGF-β1's active concentration has to be precise: 1–2 ng/mL TGF-β1 + 100 U/mL IL-2 → ~35–40% Foxp3⁺ from naïve CD4⁺ (C57BL/6, 72 h); 2 ng/mL TGF-β1 + 20 ng/mL IL-6 → ~25% RORγt⁺ / IL-17A⁺. If your TGF-β1 is 80% latent, you're at 0.4 ng/mL active → Foxp3 drops to 12%, and you mis-call your anti-CD3 dose-response. PRP100190's endotoxin <1 EU/μg is critical here: naïve T cells are LPS-hypersensitive (TLR4 on T cells is low but DCs in the culture aren't — endotoxin >5 EU/μg activates DCs → IL-6 surge → Th17 bias even at "Treg conditions," ruining your Foxp3%/IL-17A% split). For human Treg GMP work, PRP100190's low endotoxin also passes the "no pyrogen" bar for ex vivo clinical-grade cultures.
- NMuMG / AML12 EMT (The Snail/Slug, E-cad↓, Vimentin↑ Assay)
NMuMG (mouse mammary gland, immortalized, often used with or without PyMT Ras) is the standard TGF-β1 EMT readout: 2–5 ng/mL TGF-β1 (acid-activated) → 48–72 h → E-cadherin WB/IHC ↓ 60%, vimentin ↑ 3×, Snail/Slug qPCR ↑ 5–8×, cell morphology spindle-shift. The "acid-activated" step is twice as important here because EMT papers often use TGF-β1 + EGF or TGF-β1 + Wnt3a combinations — if your TGF-β1 baseline is 20% latent, the synergy ratio with EGF/Wnt shifts, and your "Alk5i rescued morphology" claim becomes unreproducible. PRP100190's Mv1Lu EC₅₀ validation guarantees the active dimer is there; just don't skip the HCl step or you'll wonder why your "5 ng/mL" looks like someone else's "1 ng/mL."
- CAF / TME Models & 3D Organoid Crosstalk
Cancer-associated fibroblasts (CAF) in PDAC (Panc02, KPC) and TNBC (4T1, MDA-MB-231 co-culture) secrete TGF-β1 into the TME, driving desmoplasia, gemcitabine resistance, and EMT-metastasis. For in vitro CAF-conditioned medium transfers: you supplement CAF medium with 5–10 ng/mL PRP100190 (acid-activated) to mimic TME-high TGF-β1, then treat with Alk5i (SB525334 5 μM) or anti-TGF-β1 to show "TGF-β1 drop → E-cad recovery in tumor organoids." For 3D gut/liver organoid-TME co-cultures, PRP100190's low endotoxin prevents "LPS-from-TGF-β1" activating stromal TLR4 and confounding your organoid viability reads. A neat trick: run TGF-β1 dose-response (0.5 / 1 / 2 / 5 ng/mL) on patient-derived organoid drug screen — PRP100190's batch CV <8% on Mv1Lu EC₅₀ means your dose-response IC₅₀ for Alk5i will reproduce across 3 organoid lines, which is what reviewers ask for in TME papers.
- Pulmonary / Renal Fibrosis Explant Counter-Screens
Bleomycin lung explants (C57BL/6, 0.5 U IT) → day 7 lung homogenate → you want to add back TGF-β1 ex vivo to prime fibroblast-myofibroblast transition in precision-cut lung slices (PCLS) → PRP100190 2 ng/mL + 1% FBS, 48 h → α-SMA, collagen I, p-Smad2/3. Or UUO kidney: renal fibroblast primary culture + 5 ng/mL PRP100190 → FN/Col3a1 qPCR. The key here is acid-activation + carrier BSA in the reconstitution: TGF-β1 adsorbs to polypropylene tube walls at <10 ng/mL concentrations (you lose 30–50% if you dilute into PBS without 0.1% BSA). PRP100190's formulation (or the reconstitution protocol in the manual) accounts for this — most commodity vials don't mention it, and your "2 ng/mL" well ends up at 0.8 ng/mL.
Quick Optimization Notes (TGF-β1-Specific, Not Generic Cytokine)
• Acid-activation is non-negotiable: PRP100190 ships LAP-clamped. Reconstitute lyophilized powder in 4 mM HCl + 0.1% BSA (carrier prevents adsorption), then for each experiment do the working-stock activation: dilute to desired concentration in 1 N HCl, incubate RT 10–15 min, neutralize 1:1 with 1 N NaOH / HEPES, then add to culture medium (medium's bicarbonate + serum will buffer to pH 7.2–7.4). Skip this and you're at ~10% active.
• Carrier BSA matters at low dose: TGF-β1 adsorbs to tube walls below 10 ng/mL. For 0.5–2 ng/mL working concentrations (Treg, Mv1Lu), keep a 100× acid-activated stock in 4 mM HCl + 0.1% BSA, dilute 1:100 into medium → final BSA <0.001%, won't affect cells.
• Storage & freeze-thaw: -20°C, aliquot into 10–20 μL single-use stocks after acid-activation (e.g., 100× stock = 1 μg/mL in 4 mM HCl + 0.1% BSA, -20°C). Each freeze-thaw loses ~10% activity (LAP can re-clamp partially if pH fluctuates, or dimer can dissociate). Avoid >2 freeze-thaw.
• Counter-screen control: Always run SB431542 (10 μM) or SB525334 (5 μM) alongside TGF-β1-treated wells — if p-Smad2/α-SMA/Foxp3 doesn't reverse ≥70%, your readout has a non-TGF-β1 component (likely endotoxin TLR4, or your "TGF-β1" had latent-only + some aggregated inactive that's actually acting as a decoy — PRP100190's endotoxin control + bioactivity QC prevents this, but still good practice).
• Don't heat-inactivate serum with TGF-β1 in it: 56°C 30 min destroys ~40% of active TGF-β1 dimer (disulfide perturbation). Use charcoal-stripped serum if you need low-endogenous-TGF, not heat-inactivated.
The Bottom Line
TGF-β1 is the 112-aa (~25 kDa non-reducing dimer, ~12.5 kDa reducing monomer) Jekyll-Hyde cytokine that drives fibrosis (liver, lung, kidney, skin), Treg polarization, EMT, and the cancer "early suppressor / late prometastatic" paradox — but >90% of it is LAP-clamped latent in any recombinant preparation, and your "10 ng/mL" spike is only ~1 ng/mL active if you skip acid-activation or bought a vial that wasn't batch-validated. The Recombinant Human TGF-β1 Protein (PRP100190) from Abbkine gives you the human mature dimer (LAP-clamped SLC, physiological form) with ≥95% purity, endotoxin <1 EU/μg, and Mv1Lu proliferation-inhibition EC₅₀ ≤ 0.5 ng/mL — meaning your LX-2 α-SMA, naïve CD4⁺ Foxp3%, and NMuMG EMT reads are driven by active dimer, not latent background. Whether you're running a CCl₄ HSC timecourse, a Treg/Th17 polarization plate, or a CAF-organoid TME screen, it's the TGF-β1 reagent that doesn't make you wonder which fraction is actually working.
Product Reference: PRP100190 – Recombinant Human TGF-β1 Protein
Learn more and order: https://www.abbkine.com/product/human-tgf-%ce%b21-protein-prp100190/
(For Research Use Only; not for diagnostic procedures in humans.)





